|
|
|
SYSTEMATIC PALEONTOLOGY
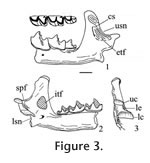
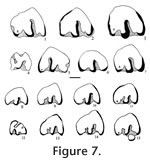
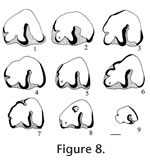
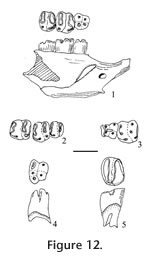
Order INSECTIVORA Cuvier, 1817 Type species. - Paranotiosorex panacaensis new species. Included species. Type species only. Diagnosis. Size is similar to Notiosorex crawfordi with dentition 1-4-3/1-2-3 (incisor-antemolars-molars). The upper incisor is non-bifid. P4, M1, and M2 have strong posterior emargination of the basal outline. The lower incisor has one low scalloped cuspule. The entoconid on m1 and m2 are small, well separated from the metaconid with a low entoconid crest. The talonid of m3 is reduced with a low cristid obliqua, joining a low, transversely elongated cusp to the posterior side of the protoconid-metaconid wall below the media notch, in contrast to the crescentic basin of Notiosorex crawfordi. The mandibular articulation has a narrow interarticular area between the two condyles with a distinct lingual emargination relative to the condyles of Sorex. The upper condyle is oval, and the lower condyle is elongate and offset lingually from the plane of the lower sigmoid notch but without a groove. There is light pigmentation on the tips of the P4, M1, lower incisor, lower antemolars, m1, and m2. Differential diagnosis. Paranotiosorex n. gen. differs from Notiosorex by its oval upper condyle on the mandibular articulation and less lingual offset of the lower condyle. It differs from Megasorex by its smaller size as well as the two differences for Notiosorex and from all other shrews by its Neomyini tribal characters. Etymology. Para, close to, implying morphologically close to Notiosorex. Occurrence. Known only from the Panaca l.f., southeastern Nevada.
Paranotiosorex panacaensis new species
Etymology. panaca, name of the town near type locality; ensis, Latin "of place." Diagnosis. as of the genus. Material examined. UALP loc. 8197: 21218, I; 21219, L maxillary with P4; 21220, L maxillary with M1-M2; 21221, R maxillary with P4-M1; 21222, R dentary with m2-m3; 21223, mandible with i-m2; 21225, L dentary with m1-m3 and complete posterior area; 21431, RM1; 23381, dentary with i-m2. UALP loc. 9601: 22855, L dentary with m2-m3. UALP loc. 9602: 22859, R dentary with two antemolars (including p4) and m1; 22860, R dentary with incisor, p4 and m1; 22861, R dentary with m3; 22862, Left maxillary with M1-M2; 22864, LM1 or M2; 22894, L dentary with m1-m3 and complete posterior area. UALP loc. 9620: 22873, m1; 22874, m1; 22875, Lm2. UALP loc. 9702: 22890, L maxillary with I-P4; 22891, L maxillary with M1-M2; 22892, LM1; 22893, I; 22850, R dentary with posterior area and m2-m3; 22886, Rm1; 22887, Rm2; 22889, i. Description. A maxillary fragment (UALP 22890) with incisor and four antemolar teeth indicates that the upper dentition of this species is 1-4-3. The incisor is falciform and bicuspid. Its anterior cusp is non-bifid, and the posterior cusp is very low and flat. A thin basal cingulum is present on the labial side but is absent on the lingual side. No pigmentation is seen on the three available incisors. The first three upper antemolars are unicuspid and minute, with well-developed cingula on both labial and lingual sides. The anterior two antemolars are subequal in size, whereas the posterior antemolar is slightly smaller. No pigmentation is seen on the three teeth. The P4 is slightly smaller than the M1 and appears trapezoidal in occlusal view. The alveolus beneath this broken P4 suggests that it had strong posterior emargination of the basal outline and a broadly curving hypoconal flange. The protocone of the P4 is small but distinct on the anterior border. The parastyle is small, joining the low anterior cingulum that connects with the protocone lingually. The paracone is the largest cusp, separated from the metacone by a broad notch. A low, narrow cingulum is present at the posterior side of the metacone. The hypocone is slightly lingual to the protocone. Very light pigmentation is seen on the crown of the P4 below the notch between the paracone and metacone. M1 is wider than long, with strong emargination of the posterior basal outline. A W-shaped ectoloph is well developed, with the posterior V distinctively larger, higher, and more elongated posteriorly relative to the anterior V. The metacone is larger and higher than the paracone, and both are much higher than the protocone. The parastyle and metastyle are prominent whereas the mesostyle is slightly less developed. The hypocone is small, low, close and slightly lingual relative to the protocone. The hypoconal flange is elongated posteriorly and slightly higher anteriorly. A narrow and low posterior cingulum is present. No pigmentation is seen on two specimens, whereas light pigmentation is seen on the ridge of the ectoloph of one specimen (UALP 22892). M2 is similar to M1 except for the following characters: smaller, anterior and posterior Vs subequal on the W-shaped ectoloph, less strong emargination of the posterior basal outline, and less developed hypoconal flange. No pigmentation is seen on the two specimens. Two mandibles from UALP loc. 9702 and one from UALP loc. 9602 preserve a complete coronoid process and condyles. The lingual condylar emargination is open, without a filled bony plate (as seen in the Blarinini). Consequently, the two condyles are separated by a narrow interarticular area. The upper condyle is oval, and the lower condyle is elongated, offset lingually from the plane of the lower sigmoid notch to a greater extent than in Blarini or Soricini, but no grooves are developed as in other species of Neomyini. The coronoid process is slightly deflected labially, and the tip of the coronoid process is slightly curved anteriorly as in Notiosorex. A coronoid spicule is prominent (Figure 3.1). The superior pterygoid fossa is shallowly basined. A pterygoid spicule is absent, but a tiny pterygoid boss can be seen. The internal temporal fossa is subtriangular, and the external temporal fossa is very shallow with the ventral margin below the level of the upper sigmoid notch. The mental foramen is located below m1, slightly anterior to the hypoconid or between the protoconid and hypoconid. UALP 22859 indicates the lower dentition of P. panacaensis n. gen and sp. is 1-2-3. The lower incisor on this specimen is broken, but UALP 22889 shows that the lower incisor is procumbent and elongated, with one low cuspule on the cutting edge. The first antemolar is unicuspid, smaller than the second antemolar (p4). The p4 has a characteristic Soricinae pattern, with an L-shaped posterolabial crest and a posterolingual basin. No pigmentation is seen on the antemolars of UALP 22859. However, UALP 23381 has light pigmentation on the tips of the incisor and antemolars. The trigonid and talonid of m1 are subequal anteroposteriorly, with the trigonid distinctly higher than the talonid. The paraconid is anterior to the protoconid and metaconid. It is also lower and farther from the protoconid than the metaconid. In the talonid, the hypoconid is high and prominent; a small but distinct entoconid is present on the lingual border, slightly anterior to the hypolophid and well separated from the metaconid. The entoconid is low and can be detected most securely from the lingual side (Figure 3.1-3.2). The anterior cingulum and the cingulum labial to the hypoconid are slightly more prominent than the posterior cingulum and the labial cingulum beneath the protoconid. UALP 22894 is exceptional in that the cingulum is uniform and better developed. Light pigmentation is seen only on one m1, UALP 23381. The m2 is similar to m1 except that it is smaller, and the paraconid and metaconid are subequal in length. Light pigmentation is seen only on one m2, UALP 23381. The m3 is much smaller than m1 or m2. Compared to m2, the protoconid and metaconid are closer, and the metaconid is slightly smaller than the paraconid. The talonid of m3 is reduced to a short and narrow basin, with a minute hypoconid and a low cristid obliqua joining the posterior wall of the protoconid-metaconid below the median notch. Reduction of the talonid is similar to that of Notiosorex crawfordi. Anterior and labial cingula are moderately developed. Discussion. The occlusal pattern of p4 and the deep, pocketed internal temporal fossa exclude P. panacaensis n. gen and sp. from the subfamily Heterosoricinae. The lingual emargination of the mandibular articulation indicates that P. panacaensis n. gen and sp. is not a member of the subfamilies Limnoecinae or Crocidurinae. All of these characters strongly suggest that this shrew belongs to the subfamily Soricinae. Strong emargination of the posterior basal outline on P4, M1, and M2 of P. panacaensis n. gen and sp. is similar to that seen in the tribe Soricini; however, the following diagnostic characters of Soricini (Repenning 1967) are not found in P. panacaensis n. gen and sp.: 1) mandibular condyles are separated, the interarticular area is broad with no conspicuous lingual emargination; 2) lower condyle is not offset lingually beyond the plane of the mandible at the lower sigmoid notch; 3) superior pterygoid fossa of the mandible is a shallow pit rather than a deep basin; 4) internal temporal fossa is large and triangular; 5) coronoid process is not deflected labially; 6) m1 has a strong entoconid crest; and 7) the upper dentition is usually 1-6-3. P. panacaensis n. gen and sp. has the following characters that are similar to those of the tribe Blarini: 1) mandibular condyles are widely separated, and the lower condyle is offset lingually beyond the lower sigmoid notch; 2) internal temporal fossa is small and oval; 3) coronoid process is slightly deflected labially; 4) no spicule is developed in the superior pterygoid fossa; and 5) m1 and m2 have no (or a very low) entoconid crest. However, members of the tribe Blarini usually have a bony plate filling the lingual emargination of the mandibular articulation so that the interarticular area looks broad, and most species of this tribe lack emargination of the posterior basal line on P4, M1, and M2, as seen in Blarina and Paracryptotis. However, some species of Cryptotis (e.g., Cryptotis parva) do not have a bony plate in the interarticular area, and the upper molars have moderate emargination. But Cryptotis is more primitive than P. panacaensis n. gen and sp. in that it retains five antemolar teeth in the upper dentition, whereas Paranotiosorex n. gen. has only four. The lingual emargination of interarticular area in the mandible, lack of bony plate filling the interarticular area, lingual offset of the lower condyle, low external temporal fossa, and the 1-4-3 upper dentition suggest that P. panacaensis n. gen and sp. belongs in the tribe Neomyini. Only two genera of Neomyini, Notiosorex and Megasorex, have been reported from North America. Megasorex is significantly larger than Notiosorex, and P. panacaensis n. gen and sp. is similar to Notiosorex crawfordi (living species) in all characters except the shape of the upper mandibular condyle. P. panacaensis n. gen and sp. has an oval upper condyle, whereas N. crawfordi has a triangular upper condyle. Also, the offset of the lower condyle from the lower sigmoid notch is stronger in N. crawfordi where a distinct groove is seen between the condyle and notch. Two fossil species of Notiosorex (N. jacksoni Hibbard 1950 and N. repenningi Lindsay and Jacobs 1985) have been reported in North America. N. jacksoni is from the early Blancan Fox Canyon local fauna (Hibbard 1950) of Kansas and the Beck Ranch local fauna (Dalquest 1978) of Texas. Measurements of P. panacaensis n. gen and sp. are smaller than those of the holotype and paratype of N. jacksoni given by Hibbard (1950). According to Hibbard (1950), there is no posterior cingulum on P4, M1, or M2 of N. jacksoni, and this was the primary reason he erected a new species of Notiosorex. The entoconid and entoconid crest on lower molars and the talonid on m3 of N. jacksoni were not described. A lower jaw collected later from the Fox Canyon fauna (Hibbard 1953, figure 4.3B) shows a triangular upper condyle of the mandible in N. jacksoni. The other fossil species, N. repenningi, from the early Blancan Concha fauna of Mexico (Lindsay and Jacobs 1985) is much larger than P. panacaensis n. gen and sp. N. repenningi has a triangular upper mandibular condyle, a small but distinct entoconid, and a low entoconid crest on m1 and m2. The talonid of m3 is reduced to a posterior distinct entoconid and a low entoconid ridge on the lingual side. The labial cingula on the lower molars of N. repenningi are very deep. P. panacaensis n. gen and sp. can be distinguished from the three species in Notiosorex by the shape of the upper mandibular condyle. Based on this character, as well as those discussed above, P. panacaensis n. gen and sp. is considered a new species in a new genus. Some Eurasian Neomyini, such as Nectogale and Soriculus, have an oval upper mandibular condyle. However, their dental morphology differs from P. panacaensis n. gen and sp. in that they have: 1) bifid upper incisors; 2) accessory cuspids on p4 and lower molars in Nectogale; 3) a reduced labial cingulum on lower molars in Nectogale; and 4) upper dental formula 1-5-3 in Soriculus. Tedford (1961) suggested that Hesperosorex from the middle Clarendonian Ricardo local fauna of California might be ancestral to Notiosorex. Hesperosorex is more primitive in having lower crown height, retaining the entoconid on m3, and having less reduction of m3 relative to m1. The upper mandibular condyle of Hesperosorex is oval, and the lower condyle has an extreme lingual offset, resulting in a groove between the condyle and the lower sigmoid notch. There is no lingual emargination on the interarticular area in Hesperosorex, in contrast to Notiosorex and Paranotiosorex n. gen. Because Hesperosorex already has a greater offset of the lower condyle, it is probably not the direct ancestor of Paranotiosorex n. gen. There is much more intraspecific variation in dental morphology of shrews within the recent specimens of N. crawfordi in the mammal collection at the University of Arizona than anticipated. Of the 11 specimens examined, eight have a small entoconid on the m1 and m2, and the entoconid crest is low. But, the other three specimens have a large entoconid with a distinctively high crest. The former characters are the same as those of P. panacaensis n. gen and sp., and the latter are identical to the diagnosis of N. crawfordi given by Repenning (1967) who compared four recent specimens from Texas and California in the Museum of Vertebrate Zoology at the University of California, Berkeley. There are three possible explanations for this phenomenon: 1) misidentification of the eight specimens in the University of Arizona collection; 2) sexual dimorphism; and 3) intraspecific variation. The UALP specimens are identified correctly; examination of the mandibular structure of the eight specimens indicated that the condyles are identical. Thus they can belong only to the tribe Neomyini. The only other species of this tribe living in North America is Megasorex gigas, which is significantly larger with a slight posterior emargination on p4, and a less reduced talonid on m3. All of these characters eliminate the possibility that the Notiosorex specimens were misidentified specimens of Megasorex. There is a single species of Notiosorex, N. crawfordi, presently recognized in North America. Therefore, the possibility of misidentification of N. crawfordi is ruled out, unless an unnamed species of Notiosorex is hiding in North America. All three of the Notiosorex specimens with a strong entoconid and high entoconid crest examined are female, but females also occur among the other eight specimens, which argues against sexual dimorphism as an explanation for the morphological variation. Therefore, it seems likely that the variation in dental morphology seen among living specimens of N. crawfordi results from intraspecific variation strongly suggesting that caution is needed when dental morphology is used to identify shrews. Dalquest (1972) erected a new genus and species, Beckiasorex hibbardi, from the Beck Ranch local fauna of Texas. Beckiasorex is similar in size to N. crawfordi, and has a mandibular articulation identical to that of Notiosorex. Dalquest (1972) stated that B. hibbardi most closely resembles N. jacksoni except for its smaller size, reduced entoconid, and low entoconid crest. It is very likely that B. hibbardi represents a new species of Notiosorex rather than a new genus. Occurrence. Known only from the Panaca l.f., southeastern Nevada.
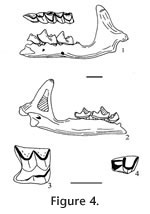
?Paranotiosorex sp. Material examined. UALP loc. 9504: 22853, R maxillary with P4-M2; 22854, R dentary with i plus p4-m3; 22867, R dentary with m2-m3; 22869, LM2; 22870, L maxillary with last two antemolar teeth and alveoli of incisor and anterior two antemolar teeth; 22871, I; 22872, i. Description. The isolated upper incisor (UALP 22871) is considered the same species as the other specimens from this locality. It is falciform and bicuspid, and the anterior cusp is non-bifid. A thin cingulum is present on its labial side of the tooth. Dark pigmentation is seen on the tip of the anterior and posterior cusps. A left maxillary (UALP 22870) indicates that the upper dentition of this shrew is 1-4-3. The third antemolar tooth is unicuspid and tiny, slightly smaller than the third antemolar of Paranotiosorex panacaensis n. gen and sp. The P4, M1, and M2 are all similar to those of P. panacaensis except for their slightly smaller size and presence of dark pigmentation on the tips of the paracone and metacone. The posterior part of the mandible on UALP 22854 is broken, but lingual emargination of the interarticular area is preserved, and the lower condyle has the same lingual offset as seen in P. panacaensis n. gen and sp. The mental foramen is located beneath the m1, slightly anterior to the hypoconid. The lower dentition is 1-2-3, as in P. panacaensis n. gen and sp. The p4 is unicuspid, with occlusal features characteristic of the Soricinae. The m1 has a high trigonid and slightly smaller talonid. The entoconid is small but distinct relative to the metaconid; the entoconid crest is short and very low. Labial cingulum is more developed than in P. panacaensis n. gen and sp. The m2 is similar to m1 except for its smaller size. The m3 is reduced in both the trigonid and talonid. The talonid is confined to a crescentic basin with a small posterior cuspule. Dark pigmentation is present on the tip of p4 and the protoconid of m1. Light pigmentation is seen on the protoconid of m2. Discussion. The following features indicate that this shrew belongs in the tribe Neomyini: 1) lingual emargination of the interarticular area in the mandible; 2) lingual offset of the lower mandibular condyle; 3) small entoconid and low entoconid crest; and 4) upper dentition of 1-4-3. The dental morphology of this taxon is similar to that of Notiosorex and Paranotiosorex n. gen. It differs from P. panacaensis n. gen. and sp. in having dark pigmentation, and the talonid of m3 lacks a distinct cusp. It may represent a different species of Paranotiosorex n. gen., or a different genus (the diagnostic upper mandibular condyle is absent). Occurrence. Known only from the Panaca l.f., southeastern Nevada.
Tribe SORICINI
Fischer de Waldheim, 1817
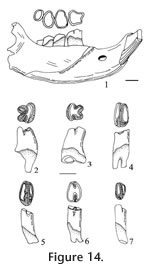
Description. This shrew is quite small. Occlusal outline of M1 is rectangular, with moderate emargination of the posterior basal margin. The posterior V is larger and slightly higher than the anterior V on the W-shaped ectoloph. The hypocone is small and slightly lingual relative to the protocone. The hypoconal flange is flat, without curving upward distally, and a posterior cingulum is present. Pigmentation cannot be detected securely because of the overall dark brown tooth color, although the tips of the paracone and metacone appear to be darker, possibly caused by pigmentation. The posterior part of the mandible (UALP 22848) is broken but the small pit of the superior pterygoid fossa is preserved, indicating a lingual emargination of the interarticular area. The lower condyle is oval, extending labially to the lower sigmoid notch. The coronoid process is not deflected labially, and it expands distally with a small external temporal boss. The external temporal fossa extends to about the level of the upper sigmoid notch. The internal temporal fossa is large and triangular. The mental foramen is beneath the m1, below the notch between the paraconid and protoconid. The p4 is unicuspid, with an L-shaped crest and a large, deep posterolingual basin. The p4 has a prominent posterolingual cingulum. The trigonid of m1 is longer than the talonid, with the metaconid slightly higher than the entoconid, and the entoconid higher than the paraconid. The entoconid is posterior to the metaconid with a high entoconid crest. The labial cingulum on m1 is thin beneath the protoconid. The lingual cingulum is well developed. The m2 is similar to m1 but smaller and with subequal height of the entoconid and paraconid. The labial cingulum is present beneath the protoconid, and the lingual cingulum is well developed. The m3 is the smallest tooth. The talonid is smaller than the trigonid, with a V-shaped ridge on the distinct sublabial hypoconid. A labial cingulum is well developed on m3. No pigmentation can be detected on the lower cheek teeth due to the dark tooth color, although the tips of cusps on p4, m1, and m2 are more heavily colored, which may be caused by pigmentation. Discussion. These specimens are assigned to the genus Sorex because of the following characters: 1) occlusal morphology of the p4 is like that seen in the subfamily Soricinae; 2) mandibular condyles have morphology characteristic of the tribe Soricini; 3) mental foramen is below m1, located anterior to the protoconid; 4) m1 has a high entoconid and well-developed entoconid crest; 5) m3 is slightly reduced and has a basined heel; and 6) posterior outline of M1 has a slight but distinct emargination. Eight species of Sorex are recognized from North America during the Pliocene and early Pleistocene (Hibbard 1937a, 1953, 1956; Hibbard and Bjork 1971). In many of the published descriptions of these species the morphology of the mandibular condyles and dentition are incomplete, making species identifications difficult. Repenning (1967) questioned the generic and tribal assignment of S. dixonensis Hibbard 1956 and S. leahyi Hibbard 1956 because these species display mandibular condyles characteristic of the tribe Neomyini (see Hibbard 1956, figures 3-4). The Panaca material is most similar to Sorex meltoni Hibbard and Bjork (1971) from the Hagerman local fauna. S. meltoni differs from S. taylori Hibbard 1937a in having a more reduced m3 and less developed labial cingulum on m1; from S. hagermanensis Hibbard and Bjork (1971), in smaller and lacking the entoconid on m3; from S. powersi Hibbard and Bjork (1971), in being smaller and having a higher entoconid crest on m1 and m2 (the entoconid crest of S. powersi was not described, but the entoconid crest seen in the illustration [Hibbard and Bjork 1971, figures 4.1C, D] is low); from S. sandersi Hibbard (1956) in being smaller; and from S. rexroadensis Hibbard (1953) by having a different internal temporal fossa. Range. Early and middle Blancan NALMA. Occurrence. Known from the Panaca l.f. Extended range includes the Hagerman local fauna, Glenns Ferry Formation, Idaho.
Order LAGOMORPHA Brandt, 1855 Virtually all fossil leporid systematics is based on the p3 and P2, therefore only these teeth were selected for detailed study. The deciduous premolars change both in occlusal size and enamel pattern, so these teeth were also excluded from the study.
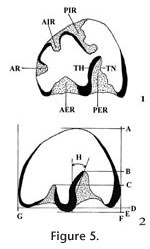
The procedure for study of leporids follows that of White (1987). In this method the maximum length and width of each p3 is measured with a dial caliper calibrated to 0.01 mm. The occlusal enamel pattern is then drawn on an index card using a microscope and camera lucida. Measurements of other features, including angles, are taken from the drawings. Abbreviations for the measurements, developed by White (1987), are shown in Figure 5.
Subfamily ARCHAEOLAGINAE Dice, 1929
Description. Hypolagus edensis is a small leporine. The p3 of H. edensis has no AR, and its AER is shallow with a relatively smooth outline. On one immature specimen (UALP 21520) a deeply incised AIR is connected to the PER, but near the base of the crown the AIR is absent and the AER is deeply incised. In the sample of 52 p3s, the TH is straight (40.4%), weakly sigmoid (32.7%), or sigmoid (26.9%) and TN is folded in 28.8% of the sample. Size of p3, along with depth ratios of AER and PER relative to width of the tooth, and deflection of PER are given in Table 2. Discussion. The p3 of H. edensis is distinguished from that of all other species of Hypolagus by a deeply incised AER. H. edensis differs from Pewelagus dawsonae, another small leporid species in the Panaca l.f., by having a relatively smooth outline of the PER. Range. Late Hemphillian and early to middle Blancan NALMAs. Occurrence. UALP loc. 8197 in the Limestone Corner section, UALP locs. 9504, 9601, and 9602 in the Rodent Hill section, UALP loc. 9702 in the Rodent Ravine section of the Panaca l.f., southeastern Nevada. The extended range includes the late Hemphillian NALMA of Arizona and California and the Blancan NALMA of California, Idaho, Texas, and Washington (White 1987; White and Morgan 1995).
Hypolagus tedfordi
White, 1987 Material examined. Limestone Corner F:AM loc.: F:AM 108711, Rp3, UALP loc. 9601: 22188, Lp3, UALP loc. 9602: 22240, Lp3, UALP loc. 9621: 22393, Rp3. Description. H. tedfordi is a small leporine whose p3 has a triangular cross section. In occlusal view, the p3 PER and AER are both shallow, PER is weakly deflected posteriorly, and the TH is sigmoid in 2 of 4 specimens. The TN is unfolded in all specimens, and the AER is not crenulated. Discussion. H. tedfordi differs from other known species of Hypolagus except H. fontinalis in its markedly shallow (weakly incised) PER. It is distinguished from H. fontinalis by its significantly smaller size (White 1987). White (1987) also mentioned that H. tedfordi usually has an anteriorly deflected PER, whereas H. fontinalis has a posteriorly deflected PER. In the three p3s assigned to H. tedfordi from the Panaca l.f., each PER is weakly deflected posteriorly. Thus, in H. tedfordi there is some variation in the deflection of the PER. H. tedfordi was found only in the Clarendonia and Hemphillian faunas, previously. The occurrence of H. tedfordi in the Panaca l.f. extends its geological range into early Blancan NALMA. Range. Clarendonian to early Blancan NALMAs. Occurrence. F:AM loc. in the Limestone Corner section, UALP locs. 9601 and 9602 in the Rodent Hill section, UALP loc. 9621 in the Brown Bone section of the Panaca l.f.., southeastern Nevada. The extended range includes late Clarendonian NALMA of California and the Hemphillian NALMA of Arizona and California (White 1987).
Hypolagus cf. H. ringoldensis
Gustafson, 1978
Description. Hypolagus cf. H. ringoldensis is a relatively large leporid with variable development of the AR; five p3 specimens in the sample of 10 from the Panaca Fm. have a thin AR that is very shallow but filled with cementum; TH is weakly sigmoid in all specimens and crenulation is absent on the enamel wall of AER. Discussion. This taxon resembles Hypolagus ringoldensis because of its large size and variable development of the AR. This taxon is questionably referred to H. ringoldensis because it expands the known characteristics of that species; e.g., the TH is weakly sigmoid, and development of internal reentrants is variable. It may represent a new species, but the sample is too small and the morphology too variable to identify consistent characters. Hypolagus cf. H. ringoldensis differs from most other leporids recovered from the Panaca l.f. by its larger size. It differs from H. regalis in the absence of a strongly anteriorly deflected PER, and the AER is not as deeply incised. Hypolagus cf. H. ringoldensis differs from H. oregonensis, known only from the late Hemphillian of Oregon (White 1987), in larger size and TH is not usually sigmoid. Range. Late Hemphillian and early Blancan NALMAs. Occurrence. UALP loc. 8197 and F:AM loc in the Limestone Corner section, UALP locs. 9601 and 9602 in the Rodent Hill section, UALP locs. 9619 and 9702 in the Rodent Ravine section, and USLP locs 9620 and 9621 in the Brown Bone Bed section of the Panaca l.f., southeastern Nevada. The extended range of H. ringoldensis includes the Hemphillian NALMA of Nebraska, Florida, and Arizona, the Blancan NALMA of Texas, Kansas, and Washington (White 1987).
Hypolagus gidleyi
White, 1987 Material examined. UALP loc. 9602: 22239, left p3. Description. The p3 is semicircular in cross section. Occlusal view of p3 shows that PER is more deeply incised than AER, it lacks an AR, the TH is straight and the TN and enamel wall of AER are not crenulated. Discussion. This specimen is identified as H. gidleyi because its size is smaller than H. ringoldensis and larger than H. edensis plus other small species of Hypolagus. It differs from Hypolagus regalis in lacking a strong anteriorly deflected PER; it differs from Hypolagus oregonensis in its straight TH. Size and morphology of the specimen place it with H. gidleyi. Range. Late Hemphillian to middle Blancan NALMAs. Occurrence. UALP locality 9602 in the Rodent Hill section in the Panaca l.f., southeastern Nevada. The extended range of H. gidleyi includes the Hemphillian NALMA of Texas and New Mexico, the Blancan NALMA of Idaho, Nebraska, Texas and Washington (White 1987).
Hypolagus regalis
Hibbard, 1939 Material examined. UALP loc. 9504: 22149, Rp3; UALP loc. 9601: 22193, Rp3. Description. The p3 is of medium size, with AER well incised across the occlusal surface, but less incised than the PER; the PER strongly deflected anteriorly, and the AER and TH are not crenulated. Specimen UALP 22193 is an immature tooth with AIR and PIR that do not extend to the base of the crown. Discussion. H. regalis is characterized by having a strong anterior deflection of PER on p3; it differs from H. voorhiesi in having a more deeply incised AER and with PER more strongly deflected anteriorly. H. regalis differs from H. arizonensis in being slightly larger (White 1987). Range. Late Hemphillian and early Blancan NALMAs. Occurrence. UALP locs. 9504 and 9601 in the Rodent Hill section of the Panaca l.f., southeastern Nevada. The extended range of H. regalis includes the Hemphillian NALMA (Santee l.f.) of Nebraska, the Blancan NALMA of Kansas (Rexroad and Wendell Fox l.f.s), and Texas (Beck Ranch l.f.)
Pewelagus White, 1984 Material examined. UALP loc. 8197: 21226-21232, seven isolated p3s. UALP loc. 9504: 21508-21510, three isolated p3s; 22150, p3; 22151, Rp3-p4; 22158, p3; UALP loc. 9601: 22185, p3; 22189, p3; 22190, p3; 22192, p3; 22195, p3; UALP loc. 9602: 22216, p3; 22218, L dentary with p3-m2; 22220, p3; 22223, p3; 22228-22229, two p3s; 22242-22245, four isolated p3s; 22248-22250, three isolated p3s; 22252-22253, two p3s; UALP loc. 9703: 22395, p3. UALP loc. 9619: 22323, Lp3; UALP loc. 9702: 22338, p3; 22347-22354, eight isolated p3s; 22360-22361, two p3s. UALP loc. 9620: 22394, Rp3. Description. Pewelagus is small; p3s have a triangular cross section with PER and AER deeply incised across the occlusal surface. AER has two folds (39 of 43) or more crenulations (4 of 43). TH is sigmoid (N=18), straight (N=15), or weakly sigmoid (N=10) and TN is folded in 40.5% of the sample. Dimensions and deflection of PER are given in Table 2. Discussion. Two species of Pewelagus are known, P. dawsonae and P. mexicanus; the former is distinguished from the latter by being smaller and with TN that is usually crenulated (White 1987). P. dawsonae differs from small species of Hypolagus in having two or more crenulations on AER. White (1987) had identified a p3 from the Limestone Corner F:AM locality as P. dawsonae. The localities in the Panaca Fm. that produce large samples of lagomorphs (e.g., Locs. 8197, 9504, 9601, 9602, and 9702) are dominated by P. dawsonae and H. edensis. This sympatry suggests that P. dawsonae and H. edensis probably occupied different niches. Based on its greatly enlarged tympanic bullae and narrow choanae, White (1987) suggested that P. dawsonae inhabited burrows for protection from predators. My results support that interpretation. Range. Blancan NALMA. Occurrence. UALP loc. 8197 and F.AM loc in the Limestone Corner section, UALP locs. 9504, 9601, 9602, and 9703 in the Rodent Hill section, UALP locs. 9619 and 9702 in the Rodent Ravine section, and UALP loc. 9620 in the Brown Bone Bed section of the Panaca l.f., southeastern Nevada. The extended range of Pewelagus includes the Blancan NALMA of California (Layer Cake, Vallecito Creek, and Coso Mountains l.f.s) and Texas (Red Corral and Blanco l.f.s).
Lepoides White,
1987
Description. Lepoides is significantly larger than other fossil archaeolagines. The P2 has three reentrants; the p3s are semicircular to triangular in cross section, with an AR, AER, and deeply incised PER. The PER is strongly deflected posteriorly; TH is sigmoid in 91.7% of the sample, and TN is folded in 75% of the sample (N=12). One dentary (UALP 22160) has an AIR in addition to the AR, AER, and PER. In this specimen the PER is inclined 58.7% of the occlusal width, with a slight posterior deflection, and the AER is inclined 21.7% of the occlusal width. The TN is not folded and the AER is not crenulated on this specimen. It is assigned to Lepoides based on its size and the posterior deflection of the PER. Discussion. Lepoides is distinguished from other large lagomorph species in the Panaca l.f. in having a posterior deflection of PER. Range. Late Hemphillian and early Blancan NALMAs. Occurrence. F:AM locality in the Limestone Corner section, UALP locs. 9504, 9601, 9602, and 9609 in the Rodent Hill section, and UALP locs. 9619 and 9702 in the Rodent Ravine sections of the Panaca l.f., southeastern Nevada. The extended range of Lepoides lepoides includes the Hemphillian NALMA of Nebraska (Santee l.f.; White 1987) and late Hemphillian (Silver Springs l.f.; Kelly 1998) and early Blancan NALMAs of Nevada.
Subfamily LEPORINAE Trouessart, 1880 Material examined. Limestone Corner F:AM loc.: AMNH(F:AM) 108715, R p3; UALP loc. 8197: 21241, L p3; UALP loc. 9601: 22174, L p3; UALP loc. 9602: 22217, L dentary with i, p3-m1. Description. Nekrolagus progressus is near the size of Sylvilagus floridanus and Hypolagus vetus. N. progressus differs from all other lagomorphs from the Panaca l.f. in having either a PIR or an enamel lake in the position of the PIR. The p3s of N. progressus usually with a shallow AR; the PER is deeply incised across the occlusal surface and may be slightly deflected anteriorly or posteriorly; the AER is shallow and uncrenulated; TH is smooth except for one specimen (UALP 22217) with a slightly sigmoid TH; TN is smooth except for one immature specimen (UALP 21241) in which TN is folded. One specimen (F:AM 10875) develops an enamel lake on the occlusal surface when the PIR is pinched off (Figure 5.8). Range. Late Hemphillian and early Blancan NALMAs. Occurrence. F:AM loc. and UALP loc. 8197 in the Limestone Corner section, UALP locs. 9601 and 9602 in the Rodent Hill section of the Panaca l.f., southeastern Nevada. The extended range of Nekrolagus progressus includes the Blancan NALMA of Kansas (Rexroad 3 l.f., Wendell Fox l.f., and Deer Park l.f.), Washington (White Bluffs l.f.), and Texas (Beck Ranch l.f.; White 1991).
Pronotolagus White, 1991 Material examined. UALP loc. 9504: 21511, Rp3. Description. Pronotolagus is a medium to large leporid whose p3 lacks AR and with AIR more deeply incised than PIR (when the PIR is present). The AER is shallow and wide; PER is incised less than half the occlusal width (White 1991). UALP 21511 (length is 2.1 mm, width is 1.65 mm) is slightly smaller than P. apachensis with a small but distinct AIR; the PER is deeply incised; the AER is shallow and unfolded; TH is strongly sigmoid and TN is folded. Discussion. Presence of AIR and absence of AR in UALP 21511 indicate this specimen is Pronotolagus. Size of UALP 21511 is close to that of P. apachensis but p3 of that species lacks a strongly sigmoid TH. Pronotolagus differs from Notolagus in having a less deeply incised AIR and the AIR often unites with AER in Notolagus, forming an isolated column in the anterior part of the tooth. Occurrence. UALP loc. 9504 in the Rodent Hill section in the Panaca l.f., southeastern Nevada. The extended range of Pronotolagus is late Clarendonian of California (Apache Canyon l.f.) and early Hemphillian of Nebraska (LeMoyne l.f., White 1991).
Order RODENTIA Bowdich, 1821
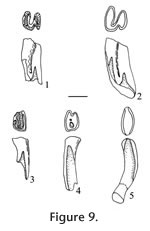
Description. Sample from the Panaca l.f. is restricted to isolated teeth; the sample is described by the type of tooth. P4: Specimen UALP 21566 (Figure 9.1) is unworn and provides a framework for loph-cusp unions; the protoloph and metaloph are connected by a prominent loph directed from the hypostyle of the metaloph to join the protoloph slightly lingual to the midline; the posterior cingulum is very low and short, about two-thirds of the length of the metaloph; dentine tracts are developed low on the sides of the protoloph with the tract on the lingual side higher than that on the labial side; dentine tracts on the metaloph cannot be determined because of breakage. UALP 21567 (Figure 9.2) is slightly worn with protoloph and metaloph joining as before; reentrant angles are V-shaped and narrow, with shallow lingual valleys and deep labial valleys; dentine tracts on the sides of the metaloph are much higher than those on the protoloph; posterior enamel surface is thin (broken on this specimen); a remnant anterior root is present.
p4: Unworn teeth have an enamel cap on the occlusal surface, with small, indistinct cusps; on worn specimens the protolophid is subcircular in outline, shorter transversely and longer anteroposteriorly than the metalophid; the two lophids are connected medially by a short prominent loph. Dentine tracts are high on the sides of the teeth, higher on the labial side, and slightly higher on the metalophid than on the protolophid. m1 and m2: Occlusal outline of m1 and m2 are similar, with the anteroposterior length slightly greater on the labial side. The metalophid has three cusps (large medial protoconid, small labial anteroposteriorly-elongated protostylid, and lingual metaconid). An anterior cingulum is well developed, joining the protostylid and the metaconid after passing anterior to the protoconid. Initially, the hypolophid is much thinner anteroposteriorly than the metalophid, with three indistinct, transversely elongated cusps. The two lophids join labially in early wear and join again (lingually) in moderate wear. The enamel base is much higher on the anterior plate than on the posterior plate, so the anterior enamel band is removed after the tooth is worn through about one-third of its height. Dentine tracts are well developed but not as narrow as seen on upper cheek teeth. Unworn and slightly worn teeth are open at the base, but worn teeth have a large single root. m3: This tooth is smaller than m1 or m2. It has a long and low anterior cingulum that continues to the lingual side of the metaconid. The hypolophid is very thin anteroposteriorly, shorter transversely, and lower than the metalophid. Cusps are indistinct, and roots are undeveloped on four available specimens. Discussion. The geomyid occlusal pattern of cheek teeth and the presence of roots on the p4s of well-worn molars indicate that this taxon is more primitive than modern Geomys, which is characterized by evergrowing teeth (Hibbard 1954a). Three species of Pliogeomys have been recorded in North America and Mexico; these are P. buisi (genotype) from the late Hemphillian Buis Ranch local fauna of Oklahoma (Hibbard 1954a), P. parvus from the middle Blancan Hagerman local fauna of Idaho (Zakrzewski 1969), and P. carranzai from the late Hemphillian Yepomera local fauna of Chihuahua (Lindsay and Jacobs 1985). P. parvus is significantly smaller than P. buisi, and the roots of p4 are not as well developed as in P. buisi (Zakrzewski 1969). P. carranzai is characterized by its very high crown, high dentine tracts, and the shallow median groove on the upper incisor. No upper incisor of a pocket gopher has been found in the Panaca l.f. Size of p4 and m2 from the Panaca l.f. are similar to P. parvus from the Hagerman l.f. (Zakrzewski 1969, table 2) although m1 is slightly larger and m3 is distinctly larger than those from the Hagerman l.f. These differences are considered within the range of intraspecific variation because the sample size of geomyids from both the Panaca and Hagerman l.f.s is small, and large variation is seen in the Idaho sample. Notice that the geomyid teeth from UALP loc. 9502 are slightly smaller than those from other Panaca sites. P. carranzai is higher crowned and larger than the Panaca geomyid. Also, dentine tracts on the sides of p4 (Lindsay and Jacobs 1985, figure 6a) of P. carranzai are much higher than those from the Panaca l.f. Martin et al. (2002) considered P. carranzai a species of Geomys. Size, crown height, and development of dentine tracts indicate that the Panaca geomyid should be assigned to P. parvus. Dalquest (1983) erected a new genus of Geomyinae, Progeomys, from the Coffee Ranch local fauna (type Hemphillian) in Texas. So far, only the genotypic species Progeomys sulcatus has been found in North America. This taxon is characterized by a strongly rooted p4 and absence of, or very incipient, development of dentine tracts on the cheek teeth. The Panaca Pliogeomys is more advanced than P. sulcatus, both in root loss and development of dentine tracts. Range. Early to middle Blancan NALMA. Occurrence. UALP locality 8197 in the Limestone Corner section, UALP locality 9502 in the Double Butte section, UALP locs. 9504. 9601, and 9602 in the Rodent Hill section, and UALP locality 9620 in the Brown Bone bed section of the Panaca l.f., southeastern Nevada. Extended range of Pliogeomys parvus includes the Hagerman local fauna of Idaho
Family HETEROMYIDAE
Allen and Chapman, 1893
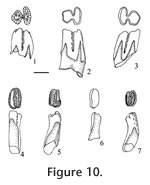
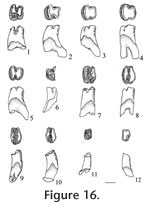
Description. Dental terminology for heteromyids follows Lindsay (1972, figure 19). Cheek teeth are brachylophodont. Upper teeth have three distinct roots (lingual, anterolabial, and posterolabial), and lower teeth have two distinct transverse roots (anterior and posterior). P4: Four cusps are present; a protocone on the protoloph, a metacone, hypocone and hypostyle on the metaloph; accessory cuspules are absent on both lophs. The protoloph and metaloph are joined medially after moderate wear; the protocone is conical, very circular. The hypocone is the largest cusp on the metaloph, the metacone and hypostyle are subequal; the hypostyle is located slightly more anterior relative to the metacone, making a smooth curved posterior outline of the metaloph; on UALP 21285 (Figure 11.3) the hypostyle is located more anteriorly, resulting in an L-shaped outline of the metaloph. Nine of the 16 P4s are slightly inflated on the lower part of the crown. The base of the enamel is lower on the protoloph. M1:Teeth are strongly bilophodont with the protoloph slightly wider transversely than the metaloph. The protoloph is relatively straight whereas the metaloph is slightly curved, especially on its posterior side. Three cusps are present on the protoloph, with the medial protocone largest and the lingual protostyle the smallest cusp; three cusps are also present on the metaloph, with the medial hypocone largest, and the lingual hypostyle the smallest. The protostyle and hypostyle are not connected until very late wear (i.e., after the other cusps are connected). The transverse valley is open lingually until wear has obliterated all of the cusps; it is open labially until very late wear. An anterior cingulum is distinct until moderate wear, connecting the protostyle to the anterolingual base of the paracone. M2: Slightly smaller and otherwise similar to the M1, except that the protoloph is distinctly wider transversely than the metaloph. Cusps are similar to those on M1. The lophs are joined lingually after early wear, forming a distinct U-shaped occlusal outline (Figure 11.4). Both anterior and posterior cingula are distinct until moderate wear. Dentary: The masseteric crest is high, as in other species of Perognathus, and the mental foramen is located below it; the average diastemal length (between the incisor and p4) is 2.73 mm (N=6); average mandibular depth below the p4 on the labial side is 2.70 mm (N=7); average alveolar length, p4-m3, is 3.90 mm (N=4). p4: Four well-developed cusps are present, two on the protolophid and two on the metalophid; accessory cuspules are absent. Labial cusps (protostylid and hypoconid) are closer than the lingual cusps (metaconid and entoconid), making an asymmetrical occlusal outline.The metalophid is slightly wider transversely than the protolophid. The transverse valley is slightly wider and deeper on the lingual side of the tooth. The two lophids lack a median ridge, with the exception of UALP 23028 and UALP 23030 (both from UALP loc. 9504) with a narrow ridge joining the protoconid and hypoconid. Union of the protolophid and metalophid occurs in moderate to late wear, joining initially on the labial side, followed (soon after) by union on the lingual side.
m2: This tooth is subrectangular in occlusal outline, transversely wider than long, with the metalophid slightly wider than the hypolophid due to reduction of the hypostylid. Cusps are similar to those on m1. The transverse valley is open on both sides, deeper on the lingual side, and lophids unite medially as in m1. m3: Size is similar to the p4; the protostylid is much reduced, and the hypostylid is absent. The metalophid is distinctly wider than the hypolophid, and the lophids are closely appressed, slightly tighter on the lingual side. The lophids are not united on any of the available specimens although they would unite medially with greater wear. The anterior cingulum is much reduced, weakly joining the protostylid and the protoconid. The transverses valley is open on both sides. Discussion. The p4 is the most diagnostic tooth of this small perognathine, with asymmetrical occlusal outline, absence of cuspules, and labial initial union of the lophids. The p4 is asymmetrical in four small fossil perognathines: P. mclaughlini, P. henryredfieldi, P. rexroadensis, and P. pearlettensis. P. henryredfieldi in late Hemphillian of Arizona (Jacobs 1977) is significantly smaller than the Panaca perognathine, and the roots of p4 are fused proximally in P. henryfieldi which distinguishes it from the Panaca perognathine. P. pearlettensis is a small species (alveolar length of p4-m3 is only 3.1 mm; Hibbard 1941a), although range of some teeth overlap those of P. mclaughlini; also, accessory cuspules may be present on p4 of P. pearlettensis. As noted by Jacobs (1977), initial union of lophids on p4 of P. pearlettensis is central rather than labial as in P. mclaughlini. P. rexroadensis is rather similar in morphology to P. mclaughlini, although "P. rexroadensis is larger and the m3 possesses a larger posterior loph for the size of the tooth than in the m3 of P. mclaughlini." (Hibbard 1950). The Panaca small perognathine is closer in size to P. mclaughlini than to P. rexroadensis, and the occlusal pattern of the Panaca small perognathine m3 is more like that of P. mclaughlini. It is therefore assigned to P. mclaughlini. Some other fossil species of Perognathus (e.g., P. gidleyi, P. stevei, P. dunklei, and P. furlongi) are similar in size to P. mclaughlini, but they all have p4s that are bilaterally symmetrical, and the initial union of p4 lophids is medial. In addition, P. gidleyi has lingual fusion of lophs in M1 and M2 (Tomida 1987); P. stevei normally has an anteroconid on p4 and frequently has a hypostylid on its p4 metalophid (Martin 1984); P. maldei has better development of an H-pattern on its lower molars (Zakrzewski 1969); and P. furlongi is more brachydont with strong inflation of the lower part of the crown, and the M1 transverse valley is open lingually until very late wear (Wood 1935; Lindsay 1972). The features above distinguish P. mclaughlini from similar size species of Perognathus. Martin et al. (2002) reviewed the features of small perognathine rodents, similar to P. mclaughlini, in their identification of a small perognathine from the Deer Park assemblages in Kansas. They assigned the Deer Park B perognathine to P. cf. gidleyi, based partly on central union of the p4 lophids although their sample included only one moderately worn p4 with the metalophid much narrower than the hypolophid. In general, small perognathine rodents are very difficult to identify and with better samples these fossil species will likely be identified more securely. Until then, identifications of small samples are often based on features that might be considered individual variation, such as central or medial union of p4 lophids. In comparison with P. mclaughlini from its type locality (Saw Rock Canyon local fauna, Kansas), specimens of P. mclaughlini from Panaca have a shorter transverse width on the lower molars, and m1 is more quadrate in occlusal outline in the Panaca specimens. These differences are considered to result from intraspecific variation, because the other dental features are so similar. Range. Late Hemphillian and early Blancan NALMAs. Occurrence. UALP locality 8197 in the Limestone Corner section, UALP locs. 9504, 9601, and 9602 in the Rodent Hill section, UALP locs. 9619 and 9702 in the Rodent Ravine section, and UALP locality 9620 in the Brown Bone Bed section of the Panaca l.f., southeastern Nevada. Extended range of Perognathus mclaughlini includes late Hemphillian NALMA of Arizona (Redington l.f., Jacobs 1977) and Oklahoma (Buis Ranch l.f., Hibbard 1954a) and early Blancan NALMA of Kansas (Saw Rock Canyon l.f., Hibbard 1949)
Oregonomys Martin, 1984 Material examined. UALP loc. 9504: 23064, left P4. Description. A large, relatively worn, brachylophodont upper premolar with protoloph elongated transversely is assigned to this genus. The protoloph is oriented obliquely relative to the metaloph, with the labial side of the protoloph placed more anteriorly than the lingual side. The metaloph has three cusps (a large medial hypocone, a labial metacone, and minute lingual hypostyle), and with the hypostyle oriented anteroposteriorly and placed slightly anterior relative to the hypocone. The transverse valley is deeper labially, partly closed lingually by the hypostyle. The posterior cingulum is well rounded, joining the hypostyle, the posterior hypocone and the metacone. Three large roots are present, fused at their bases. Discussion. The transverse elongation of the protoloph suggests that an accessory cuspule may have been present initially, erased by early wear. The size and suspected multicusped protoloph characterize this P4 as a species of Oregonomys. Three species (O. pebblespringensis, O. sargenti, and O. magnus) of Oregonomys are known. The Panaca specimen is excluded from O. sargenti whose hypostyle is not placed anteriorly, and whose union of lophs is more medial than in the Panaca P4. Size of the P4 places it in the range of measurements (see Martin 1984, tables 10 and 12) for both O. pebblespringensis (larger) and O. magnus (smaller). Occlusal pattern of the Panaca P4 is very close to that of O. magnus, although the P4 pattern of O. pebblespringensis is also possible. More specimens are needed to clarify species identification. Range. Late Hemphillian to middle Blancan NALMAs. Occurrence. UALP locality 9504 in the Rodent Hill section, Panaca l.f., southeastern Nevada. Extended range of Oregonomys includes fauna from the Dalles Formation near Arlington and McKay Reservoir in northern Oregon (Martin 1984), and the Hagerman local fauna of Idaho (Zakrzewski 1969).
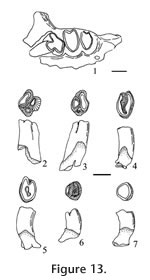
?Cupidinimus sp. Wood, 1935 Materials examined. UALP locality. 9502: 21561, left p4; 21564, left M1. UALP locality 9504: 23270, right m1. Description. Cheek teeth are mesodont, with steep crowns and deep roots. The M1 is moderately worn with cusps obliterated on the lophs; the protoloph is slightly wider transversely and straighter than the metaloph. The lophs are joined lingually by apparent fusion of the protostyle and hypostyle; the base of the protoloph is slightly lower than the base of the metaloph. The transverse valley is moderately wide and deep on the labial side. A low but broad dentine tract is present on the lingual side of the tooth. The p4 (Figure 12.4) is only slightly worn, with five distinct cusps (protoconid and protostylid on the protolophid; metaconid, hypoconid, and hypostylid on the metalophid). The protoconid and protostylid are more robust than the transversely elongate metaconid and the hypostylid; the hypoconid is smaller and medial on the metalophid. The m1 (Figure 12.5) is high crowned with a subrectangular occlusal outline, wider than long; width of the metalophid and hypolophid are subequal, each with the remnant of three cusps. The metalophid is elongated anteroposteriorly on the labial side, reflecting a well-developed anterior cingulum that would join the protostylid and the metaconid in unworn teeth. Union of the lophids occurs medially initially, in late wear, then labially; height of the labial reentrant above the base of the crown is 0.60 mm, the ratio of this height to the transverse width is 0.42, within the range of this character in Cupidinimus (0.36 to 0.67, Barnosky 1986). There is no entine tract on the sides of the tooth; base of the metalophid is markedly higher than the base of the hypolophid. There are two well-developed transversely-elongated roots. Discussion. These teeth are very similar to Prodipodomys from the Panaca Fm. based on the occlusal outline; however, these teeth are distinctly lower crowned, and the transverse valleys extend transversely wider, across the tooth, compared to the teeth of Prodipodomys. Cupidinimus is known primarily from faunas older than Blancan NALMA. Occurrence. UALP locality 9502 in the Double Butte section, and UALP locality 9504 in the Rodent Hill section of the Panaca l.f., southeastern Nevada.
Subfamily DIPODOMYINAE Coues, 1875 The classification of Korth (1994) for the Subfamily Dipodomyinae is followed except that Dipodomys minor Gidley 1922 is recognized here as Prodipodomys minor. There has been confusion among taxonomists for a number of years regarding the distinction between Dipodomys, the modern kangaroo rat, and its presumed ancestor, Prodipodomys. Gidley (1922) erected the fossil species Dipodomys minor on the basis of a mandibular fragment with p4 and the alveoli of m1-m3 from the Benson local fauna of Arizona. Gazin (1942) later removed D. minor from Dipodomys and established for it a new genus, Prodipodomys, mainly on the basis of alveoli for two well-developed roots of m1 in the holotype specimen; Gazin interpreted Prodipodomys as the primitive rooted ancestor of derived and rootless Dipodomys. This opinion was followed by Hibbard (1953), who noted that premolars of Dipodomys were often rooted in adult animals with late tooth wear, and later (Hibbard 1954b) that some extant species of kangaroo rat also have rooted molars late in life. The distinction between Dipodomys and its ancestor Prodiposomys was clarified by Zakrzewski (1981), who emended the diagnosis of both fossil genera based on crown height rather than root loss. Zakrzewski (1981) pointed out that roots of Dipodomys are generally a tubular continuum of the crown; primitive species may possess more than one root but they are generally fused into a single columnar root, also in primitive species the p4 is two rooted, and the P4 is 3-rooted. Of greater significance, Zakrzewski (1981) pointed out the crown height of teeth in adults of Dipodomys generally exceeded the length of the root, whereas the crown height in adults of Prodidodomys is approximately the same as the root length. Also, dentine tracts are well-developed, and the anterocone of P4 is lost in early wear of Dipodomys, whereas dentine tracts are absent or poorly developed, and a distinct anterocone is generally present on P4 of Prodipodomys. Characteristics of P. minor were never securely established because the taxon was based on a single specimen that lacked features of crown height and root length. A large sample of P. minor from the Panaca l.f. provides better characterization for the species. Following Zakrzewski's (1981) criteria for differentiation of Prodipodomys and Dipodomys, I conclude that it is appropriate to assign the sample of this taxon from the Panaca Fm. to Prodipodomys, as P. minor. The reasons are: 1) The dentine tracts on the sides of cheek teeth are low (<1 mm); 2) The p4s and m1s have two distinct roots; 3) Crown height of these teeth is comparable to that of P. kansensis from the Edson Quarry fauna in Kansas (Hibbard 1937b), and much lower than that of any species of Dipodomys. Height of the dentine tract on these teeth is measured relative to the base of the enamel on the labial side of the tooth. However, it should be noted that the enamel base usually has a different height on the anterior and posterior surfaces. For upper molars, the posterior enamel base is slightly higher than the anterior enamel base; the opposite pattern applies for the lower molars. Since height of a dentine tract for heteromyid teeth is usually referenced to the enamel base on the side that is higher, regardless of its location on the anterior or posterior side of the tooth, I chose the higher enamel base as the reference for the base of the dentine tract. For p4, I chose the posterior enamel base because it is higher than the anterior enamel base. In contrast, height of crown is measured relative to the lower base of the enamel for P4. Worn specimens will have lower average crown height, so height of crown should be measured only on unworn or slightly worn teeth. This procedure differs from that of Zakrzewski (1970) and Tomida (1987) who included all of the available specimens to characterize crown height in heteromyids.
Prodipodomys minor
Gidley, 1922 Emended diagnosis. A small heteromyid rodent with hypsolopodont and rooted cheek teeth. The P4 and M1 have three roots; M2 and M3 have a single root; p4 has two roots; m1 has one or two roots; m2 and m3 have a single root. Incipient dentine tracts occur on the sides of p4. Crown height of teeth is significantly lower than in Dipodomys. Materials examined. UALP locality 8197: 21406, L maxillary with P4-M1; 21407-21408, two M1s; 21409, LM2; 21411, Lp4; 21412, Rm2; 21413, m3; 21414, RP4; 21418, Lm1; 21420, m3. UALP locality 9502: 21578, Rm3; 21589, Rm3. UALP loc. 9504: 21525, P4; 21534, M1; 21536, m2; 21538-21542, five M3s; 21551, R maxillary with P4-M2; 23100, L maxillary with P4-M2; 23101, R dentary with p4-m3; 23102-23103, two P4s; 23104, R maxillary with M1-2; 23105-23108, four isolated M1s; 23109-23111, three isolated M2s; 23112, M3; 23114-23115, two M3s; 23116, Lp4-m1; 23117, p4; 23119-23120, two m1s; 23121, m2; 23122, m1; 23123-23124, two m2s; 23125, m1; 23128, m1; 23129, m3; 23131, M3; 23133, m3; 23139, M1. UALP loc. 9601: 23179, M1; 23182, Lm1; 23183-23184, two M3s; 23186, Rm2. UALP loc. 9602: 23146, Rm3; 23149-23150, two m2s; 23151, m1; 23152, m2; 23168, m1. UALP loc. 9616: 23231, RM1; 23232, LM3. UALP loc. 9620: 23241, Lp4; 23242, Rm2. UALP loc. 9621: 23247, LM3; 23248, m3. UALP loc. 9702: 23191-23192, two P4s; 23193, LM1; 23194, LM3; 23195-23196, two P4s; 23197, p4; 23198, m1; 23199, Lm2; 23200, Lm3; 23222, m1.
P4: Occlusal surface includes a narrow protoloph and a transversely wide metaloph; on unworn specimens (e.g., UALP 21414 and 23102) the protoloph has a minute accessory cuspule labial to the protocone (Figure 13.2). The metaloph has three cusps (large medial hypocone, labial metacone, and lingual hypostyle). The hypostyle is displaced anteriorly, about equidistant from the hypocone and the protocone, and remaining isolated until late wear; the hypostyle is circular on most specimens, anteroposteriorly-elongated on three specimens (UALP 21414, 23153 and 23271). The lophs are connected medially, then lingually, after pronounced wear. A low dentine tract is present on the lingual side, measured in four specimens (Table 6). Three well-developed roots are present, united basally, separate distally. M1: Occlusal surface is bilobed, transversely wider than long. The protoloph has three cusps (labial paracone, medial protocone, and lingual protostyle); the metaloph has three cusps (labial metacone, medial hypocone, and lingual hypostyle). The metaloph is transversely narrower than the protoloph and is inflated posteriorly at the hypocone, presenting a gently rounded posterior margin. The hypostyle joins the protostyle after early wear, producing a U-pattern in occlusal view. Dentine tracts are low, measurable on nine specimens (Table 6). Roots are short relative to the height of crown; 8 of 11 specimens have three roots broadly joined at the base, the lingual root is smaller than the posterolabial root and transversely-elongated anterolabial root; 2 of 11 specimens have two roots, with labial roots fused in UALP 23105, lingual and anterolabial roots fused in UALP 23193. M2: Occlusal pattern as in M1, but size is significantly smaller and transverse width of the metaloph is more reduced relative to the protoloph than on M1. The hypostyle is more reduced than on M1. Lophs are joined lingually initially; in one specimen (e.g., UALP 21409) the lophs are initially united lingually, then labially, producing an enamel islet in the center (Figure 13.5). Roots are small, more fused at the base, tending toward a single root, with a shallow groove on the labial side.
p4: Occlusal pattern is bilobed, with a lingual protoconid and labial protostylid on the protolophid, and three cusps (lingual metaconid, medial hypoconid, and labial hypostylid) on the metalophid. The metalophid is transversely wider and straighter than the protolophid. The protoconid and protostylid are elongated posteromedially, and joined medially with the metalophid in early wear. A deep anterior groove on the protolophid persists until late wear. The metaconid is the largest cusp on the metalophid, with subequal hypoconid and hypostylid. The transverse valley is wider lingually than labially, and these valleys become shallow reentrants after moderate wear, persisting almost to the base of the crown. A very low dentine tract is present on both sides of the tooth, measurable on four specimens (Table 6). Two transversely wide roots are well developed and fused proximally at the base of the crown. m1: The occlusal pattern is bilobed, with metalophid transversely wider and flexed relative to the more straight and narrow hypolophid. The largest cusp on the metalophid is the medial protoconid, with a smaller metaconid and a much smaller protostylid. An anterior cingulum joins the metaconid and the protostylid, passing anterior to the protoconid. The largest cusp on the hypolophid is the medial hypoconid, with a smaller entoconid and a much smaller and lower hypostylid. The lophids are united medially after moderate wear, forming an H-pattern. Dentine tracts are better developed on m1 than on other lower molars, and are usually present on the metalophid, measurable in 11 specimens (Table 6). The roots are fused proximally, separate distally, tending toward a relatively small, single, curved root; 8 of 12 specimens have a shallow groove on the labial side of the root. m2: The m2 is smaller than m1, with a similar occlusal pattern. The protostylid and hypostylid are both more reduced than on m1, and the lophids are joined labially at a later wear stage than the lingual union of lophids. Low dentine tracts were measured on 11 specimens (Table 6). Roots are fused proximally, separated posteriorly, tending toward a relatively small, columnar, and posteriorly curved root. Eight out of 12 specimens have a shallow groove on the labial side of the root. m3: The m3 is distinctly smaller and transversely narrower than m2. The metalophid has three cusps (metaconid, protoconid, and a much reduced protostylid). The hypolophid is lower and transversely narrower than the metalophid, it lacks a hypostylid. On unworn specimens, 2 of 3 have a low anterior cingulum, and one (UALP 21420) has a cingulum joining the protostylid and protoconid. A dentine tract is absent. Roots are fused proximally to become a long single root lacking grooves. Discussion. Size of Prodipodomys minor is similar to that of P. kansensis and P. griggsorum from Kansas; it is significantly smaller than other species of Prodipodomys and of two fossil species of Dipodomys (D. hibbardi and D. gidleyi). P. minor is considered a derived species of Prodipodomys based on fusion of the roots in both upper and lower molars. Prodipodomys kansensis from the Hemphillian Edson Quarry local fauna of Kansas (Hibbard 1937b and Zakrzewski 1970) differs from P. minor in having two well-developed roots on m1 with no dentine tracts on cheek teeth, a large mandibular foramen labial to m3, and a slight groove on the lower incisor. Prodipodomys griggsorum is slightly higher crowned than P. kansensis, but is considered more primitive than other species of Prodipodomys because of its better developed (e.g., more primitive) roots. The M1 and M2 of P. griggsorum have three well-developed roots, and the m1 has two roots. Other species of Prodipodomys from Blancan faunas are significantly larger than P. minor. In addition, P. centralis has better developed roots on M1 and M2; its p4 has accessory cuspules and a well-developed ridge between the two lophids (Hibbard 1941b, 1954b, and 1972). P. tiheni from the Borchers local fauna of Kansas (Hibbard 1943) has p4 and m1 with two roots, plus m2 and m3 are single-rooted; this is similar to roots of P. minor, however, dentine tracts on P4 and p4 of P. tiheni are slightly higher than those of P. minor (Zakrzewski 1970, table 1). P. idahoensis from the Hagerman local fauna of Idaho also has higher dentine tracts (Zakrzewski 1969, figure 5A) than P. minor. Unfortunately, the coefficient of variance (CV) for height of the dentine tracts is very high for most of the species of Prodipodomys due to the small range of height. Range. Blancan NALMA. Occurrence. UALP locality 8197 in the Limestone Corner section, UALP loc. 9502 in the Double Butte section, UALP locs. 9504, 9601, and 9602 in the Rodent Hill section, UALP locs. 9616 and 9702 in the Rodent Ravine section, and UALP locs. 9620 and 9621 in the Brown Bone Bed section, of the Panaca l.f., southeastern Nevada. Extended range of P. minor includes the Benson local fauna, San Pedro Valley, Arizona (Gidley 1922).
Prodipodomys tiheni
Hibbard, 1943
Description. Prodipodomys tiheni is larger than P. minor, sharing many morphological features with that species.
P4: Subtriangular occlusal pattern with a conical protoloph and transversely wider metaloph. The transverse occlusal outline of the protoloph increases lingually with wear. The protoloph and metaloph are broadly joined medially although the initial union may be slightly labial to the midline. The hypostyle is elongated anteroposteriorly, joining the protocone after moderate wear. Dentine tracts are low on the lingual side of tooth. Three roots are present, with the posterior roots fused at the base. M1: Bilophodont occlusal pattern with the protoloph slightly wider transversely and less inflated posteriorly than the metaloph. The protoloph has three cusps (labial paracone, medial protocone, and lingual protostyle); the metaloph has three cusps (labial metacone, medial hypocone, and lingual hypostyle. The protoloph has a gently rounded posterior margin. Lophs are joined lingually during early wear, forming a U-shape occlusal pattern. The transverse valley is deeper lingual to the midline. Dentine tracts are low on all specimens. Three roots are well-developed, with slight fusion at the base of the lingual and anterolabial roots; the anterior root is largest, and the posterolabial root is smallest. UALP 23136 has a bifurcated lingual root. M2: Occlusal pattern is similar to that of M1, but smaller and the transverse width of the metaloph is shorter than the protoloph width. The lophs are joined lingually in early wear, forming a U-shape occlusal pattern. Dentine tracts are low on all specimens (Table 6). Roots are variable with 3 of 11 having three separate roots, 4 of 11 having two roots (anterolabial and lingual roots fused), 3 of 11 having V-shaped roots, and 1 of 11 having a single root with a deep labial groove. M3: Smallest cheek tooth of upper dentition, with indistinct cusps on both lophs. The occlusal pattern is oval, becoming semicircular in late wear. The metaloph is inflated posteriorly and well rounded; the protoloph is straight, slightly wider transversely than the metaloph. Lophs are joined both lingually and labially after early wear, forming a deep enamel islet medially (Figure 15). Dentine tracts are low on 1 of 3 specimens. Two specimens have a single root, one specimen has two roots. dp4: Brachydont tooth with three transverse lophids in the occlusal pattern. The anterior lophid has the anterior cingulum lacking cusps; the protolophid and metalophid are joined medially after early wear, forming an H-pattern. p4: Bilophodont occlusal pattern with the protolophid transversely narrow relative to the metalophid. The protolophid has a shallow anterior inflection between the protoconid and protostylid that is reduced with wear, and a minute accessory cuspule is present between the protoconid and protostylid in 3 of 9 specimens (UALP 21526, 23215, and 23229). The metalophid has three cusps (lingual metaconid, medial hypoconid, and small labial hypostylid). Lophids are joined medially after early wear. A broad dentine tract is low on both labial and lingual sides of the metalophid. Two transversely elongate roots are present. m1: Bilophodont occlusal pattern with the metalophid flexed and slightly wider transversely relative to the straighter hypolophid. Three cusps occur on the metaphid (a lingual metaconid, a larger and medial protoconid, and much smaller labial protostylid). The anterior cingulum joins the metaconid and protostylid, passing anterior to the protoconid. Three cusps are present on the hypolophid (a lingual entoconid, a larger medial hypoconid, and a much smaller labial hypostylid). The lophids are joined medially after early wear, forming an H-pattern. Dentine tracts are low on both sides of the tooth. Two transversally elongated roots are present, fused at the base and separate distally. m2: Occlusal pattern as in m1, but smaller. The protostylid and hypostylid are smaller and lower than in m1. Dentine tracts are low on both sides of the tooth. Roots are variable: 3 of 8 specimens have a single root with a labial groove, 3 of 8 specimens have a V-shaped root, and 2 of 8 specimens have a single root, lacking a groove. m3: Smallest cheek tooth in the lower dentition. The metalophid has two indistinct cusps (lacking the protostyle) and a weak anterior cingulum; the hypolophid is shorter, lower and transversely narrower than the protolophid, lacking the hypostylid. A dentine tract is low on 4 of 10 specimens. A single root, curved posteriorly, is present. Discussion. Cheek teeth of Prodipodomys tiheni from the Panaca l.f. are significantly larger and slightly higher crowned than those of P. minor; they also have slightly higher dentine tracts. The P4 of P. tiheni has a hypostyle that is more elongated anteroposteriorly and connects with the protocone in early wear, in contrast to P. minor whose hypostyle is isolated until later wear. The M1 and M2 of P. tiheni are similar to those of P.minor in the development of cusps and lophs. A minute accessory cuspule occurs on the protolophid of p4 in some specimens of P. tiheni; this is probably a highly variable feature, but no accessory cuspules were seen on the equivalent p4 sample of P. minor from the Panaca l.f. Tomida (1987) reported a similar small accessory cuspule on one specimen of Dipodomys hibbardi from the 111 Ranch local fauna of Arizona. The m1 and m2 of P. tiheni are very similar to those of P. minor from the Panaca l.f., but dentine tracts are significantly higher on the P. tiheni specimens. Also, roots are better developed on m1 specimens of P. tiheni, in comparison to those of P. minor. Prodipodomys tiheni is similar in size to P. centralis from the Rexroad local fauna of Kansas and P. idahoensis from the Hagerman local fauna of Idaho. Unworn p4s of P. centralis are characterized by the resemblance to p4s of the extant heteromyid Liomys (Hibbard 1941b). An additional difference between P. tiheni and P. centralis mentioned by Hibbard (1972) is that the former has a deeper depression dorsal and labial to the mandibular foramen. Zakrzewski (1981) noted higher dentine tracts in specimens of P. tiheni relative to specimens of P. centralis from the Borchers local fauna of Kansas. Similarly, the dentine tracts of P. tiheni from the Panaca l.f. are closer to those of P. tiheni than to P. centralis from The Borchers local fauna measured by Zakrzewski (1981, table 1). P. tiheni and Dipodomys hibbardi have very similar occlusal patterns on P4 and p4, but the former is more primitive in its lower crown height, lower dentinal tracts, and better-developed roots. Similarity of the occlusal pattern suggests that P. tiheni may be close to the ancestry of Dipodomys hibbardi. In the Panaca l.f., both P. tiheni and P. minor were usually found at most of the localities. Their relative abundances are similar, and there is no significant change morphologically in these samples through the stratigraphic levels. Range. Blancan NALMA. Occurrence. UALP locality 8917 in the Limestone Corner section, UALP localities 9504, 9602, and 9703 in the Rodent Hill section, UALP locs. 9616, 9619, and 9702 in the Rodent Ravine section, UALP locs. 9620 and 9621 in the Brown Bone Bed section, UALP loc. 9502 in the Double Butte section, of the Panaca l.f., southeastern Nevada. Extended range of P. tiheni includes the Borchers local fauna, Kansas (Hibbard 1941a). |
|