|
|
|
Systematic Paleontology
Order SORICOMORPHA Gregory, 1910
Genus Cryptotis Pomel, 1848
Localities: All specimens were recovered from locality RV-7702.
Diagnosis: A species of the genus Cryptotis in which the talonid of M3 is crescent shaped. The talonid is less reduced than in Cryptotis adamsi (Hibbard) or Cryptotis parva (Hay), but lacks a distinct entoconid as
observed in C. gracilis, C. endersi, and C. magna.
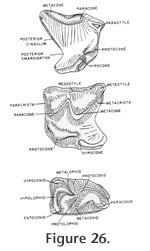
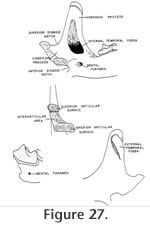
Description: The maxillaries and mandible that are assigned specimen number UCMP 317826 were found associated within a small block of matrix and show the same amount of wear. They are therefore considered to represent one individual and much of the following description is based on this specimen. Morphologic terminology utilized for the following description is illustrated in Figure 26 and Figure 27.
All lower molars are pigmented at the tips of the cusps. The lower incisor is simple with one distinct node at the anterior tip and one weak node at the posterior end of the enamel. Both nodes are lost with wear. The alveoli preserved on UCMP 317892 indicate a double rooted antemolar (P4?)
and an incisor.
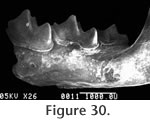
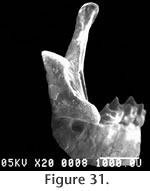
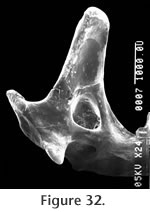
The superior and inferior articular surfaces of the mandibular condyle are separated by a relatively long and narrow interarticular area with a lingual emargination (Figure 31). The inferior articular surface is placed anterior and slightly lingual to the superior surface and is posterior to the inferior sigmoid notch (Figure 32). The inferior articular surface is roughly rectangular in shape while the superior surface is more approximately triangular. The entire condyloid process is deflected lingually. P3 is only represented by a broken base on UCMP 317826, but it would appear to be much reduced relative to P4.
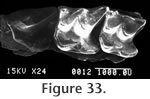
M1 is trapezoidal in shape with a strong posterior emargination (but weaker than in P4) (Figure 33). The parastyle is strong and essentially extends the posterior edge of the shearing blade of P4. The paracrista is short and perpendicular to the axis of the tooth row while the metacrista is long and sweeps posterolabially to the metastyle. The metacone is larger than the paracone. The mesostyle is placed just posterior to the labial junction of the paracone and the metacone. The protocone is well developed at the anterolingual edge of the tooth. As in P4, the hypocone is merely a slight anterior inflation on the hypoconal crest and is lingual to the posterior extension of the protocone. The hypoconal crest projects posteriorly and curves to the posterior edge of the tooth. A very weak cingulum is present anterior of the protocone while a slightly more prominent lingual cingulum extends from the base of the protocone to the base of the hypocone. A posterior cingulum is developed as in the P4.
Discussion: A number of criteria indicate that the shrew from the Warren Local Fauna is best assigned to the genus Cryptotis. The presence of an internal temporal fossa shows that it is a soricid rather than a Heterosoricine; the separation of the articular surfaces, the lingual emargination of the interarticular area, and the presence of pigmentation on the molars show that it belongs to the family Soricinae; the complete lack of an entoconid crest shows that it belongs to the tribe Blarinini and the reduced talonid of M3, the posterior emargination of P4-M2, the strong curving hypoconal crest of P4-M2, the origination of the zygomatic process opposite the metacone or metastyle of M2, and the relatively narrow interarticular area of the mandibular condyle indicate assignment of these specimens to Cryptotis. Repenning (1967) assigned a fragmentary specimen from the Hemphillian, Christmas Valley Fauna of Oregon to Cryptotis adamsi as the oldest occurrence of this genus. Re-examination of that specimen indicates that it is not Cryptotis because of a well developed entoconid crest on M1. Cryptotis sp. from the Warren Local Fauna may be the oldest known record of this genus although Wagner (1981) reported Cryptotis from the late Hemphillian Modesto Reservoir Local Fauna. All species of Cryptotis, except the questionably assigned C. meadensis Hibbard (1953), lack an entoconid crest but retain a strong entoconid. This character is diagnostic of the tribe Blarinini that Repenning (1967) suggested was derived from a primitive "sorex" form such as Antesorex in the early Miocene. He proposed a phylogeny involving the Barstovian genus Adeloblarina giving rise to both the Blarina and Cryptotis lineages. Cryptotis sp. exhibits certain primitive characters that differentiate it from younger species. Some of these characters are also observed in Adeloblarina and include: lack of labial deflection of the coronoid process, the anterior position of the mental foramen, and the slenderness of the horizontal ramus and coronoid process. Unfortunately, no M3 is known from Adeloblarina to compare with the Cryptotis sp. in which this feature is less reduced (i.e., less derived) than in younger species. The lack of a coronoid spicule and the minimal anteroposterior shortening of the talonid of M1-2 in Cryptotis sp. are two characters that appear to approach the primitive soricine condition closer than does Adeloblarina. For these reasons, Cryptotis sp. is probably not a direct descendent of Adeloblraina, however, these features do not exclude Cryptotis sp. from being ancestral to later species of Cryptotis. This evolutionary progression would require further reduction of the talonid of M3, the anteroposterior shortening of the talonids of M1-2, the posterior migration of the mental foramen, the labial deflection of the coronoid process, and the development of the coronoid spicule. All of these characters are realized in the early Blancan form Cryptotis adamsi. No upper teeth are known from Adeloblarina. The upper dentition of Cryptotis sp. is relatively primitive with the retention of P3 and the deep posterior emargination of P4-M2. These features are similar to the primitive soricine condition as seen in Antesorex. Cryptotis sp. is consistent with the suggestion by Repenning (1967) that the tribe Blarinini was derived from the primitive Soricini complex in the Miocene. The suite of characters in Cryptotis sp. contrasted with that of Adeloblarina suggests that more than one lineage attained the blarininine stage of evolution during the Miocene. Cryptotis from the Warren Local Fauna probably represents an undescribed species. Due to the limited amount of material presently available, a new name is not proposed here.
Order ERINACEOMORPHA Gregory, 1910 Material: UCMP 320133 – partial edentulous right mandible, UCMP 320134 – partial edentulous right mandible, UCMP 320135 – partial edentulous left mandible, UCMP 320597 – humerus. Localities: Specimens UCMP 320133, 320134, and 320135 - RV-7702; UCMP 320597 - RV-6834. Description and Discussion: Three mandible fragments are toothless and lack mandibular condyles. The horizontal ramus is relatively slender although only the alveoli for M2 and/or M3 are preserved. Mandibular depth beneath the anterior alveolus of M3 for each specimen is: UCMP 320133 – 2.2 mm, UCMP 320134 – 2.2 mm, and UCMP 320135 – 2.4 mm.
A
Order Lagomorpha Brandt, 1855
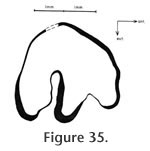
Localities: UCMP 320128 - RV-6834, UCMP 320129 and UCMP 320139 - RV-7702, and UCMP 320138 - RV-8130. Description and Discussion: The diagnostic characters used to assign this material to Hypolagus vetus Kellogg are the relatively large size of this rabbit and the morphology of P3. Both P3s (UCMP 320128 & 320138) have relatively shallow anteroexternal reentrants and deep posteroexternal reentrants (Figure 35). The occlusal patterns compare well with H. vetus described by Dawson (1958) from the type locality at Thousand Creek, Nevada, and to the emended diagnosis for this taxon provided by White (1987). The anteroposterior length of UCMP 320128 is 3.3 mm, and the transverse diameter is 3.2 mm. The size of this specimen corresponds to the largest size observed by Dawson (1958) for the topotypic material from Thousand Creek and is within the range of more than 90 samples measured by White (1987). The same dimensions for UCMP 320138 are 2.9 mm x 2.5 (+) mm at the occlusal surface, but this tooth is less worn than UCMP 320128, and the dimensions would increase significantly with wear. The anteroexternal reentrant extends approximately 24% across the occlusal surface while the posteroexternal reentrant extends 52%. This compares well with values documented by White (1987) for H. vetus. The posteroexternal reentrant is deflected slightly to the anterior. The anterior wall of both P3s has a very faint dorso-ventral groove differing from the deeper anterior reentrant in both H. oregonensis Shotwell and H. ringoldensis Gustafson. White (1987) considered Hypolagus vetus to be restricted to the Hemphillian and Blancan of the western USA and Baja, California.
Hypolagus edensis Frick
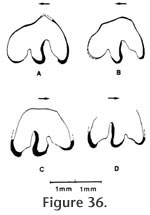
Localities: UCMP 320609 – locality RV-6834. All other specimens were collected from locality RV-7702. Description and Discussion: A second rabbit is present in the Warren Local Fauna and is readily distinguished from Hypolagus vetus by its much smaller size and by a deeper anteroexternal reentrant (Figure 36). White (1987) characterized H. edensis in his emended diagnosis as distinguishable from other species by the presence of a deep and smooth sided anteroexternal reentrant on P3. The posteroexternal reentrant of P3 in the Warren specimens extends lingually 50% of the way across the occlusal surface (n=3) consistent with the 45-59% range reported by White (1987) for this taxon. The posterior wall of this reentrant possesses a distinct anterior inflation as figured in numerous specimens of H. edensis by White (1987). The anterolabial reentrant extends 32-33% across the occlusal surface comparing well with the 24-46% range reported by White (1987). The holotype of this taxon represents an extreme in the morphologic variability with the anteroexternal reentrant being subequal in occlusal extent to the posteroexternal reentrant. v also noted that the holotype is the smallest example of this species and is from an immature individual. Both antero- and posteroexternal reentrants of the P3 of H. edensis from Warren are filled with cement. The occlusal outline of P3 is variable although the lingual wall is generally rounded. Hypolagus edensis is known from Hemphillian faunas of Arizona and California (Redington, Mt. Eden, Pinole) and the Blancan of California, Idaho, Nevada, Texas, and Washington (White 1987).
Order RODENTIA Bowdich, 1821
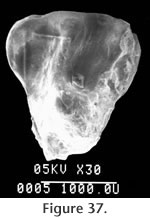
Locality: All specimens were collected from locality RV-7702. Description and Discussion: Six isolated teeth questionably are assigned to the genus Spermophilus Cuvier. A P4 (Figure 37) is triangular with a strong parastyle connected labially to the paracone. A strong protoloph connects the paracone and the protocone while a weaker metaloph connects the protocone to the metacone. A worn upper molar (M1 or 2) is approximately square with a strong protocone that joins both anterior and posterior cingula. The protoloph projects to the anterolabial edge of the protocone, but the metaloph terminates labially of the protocone. The lower dentition is represented by two M1s and two M2s. The M1s are subquadrate with partially closed trigonid basins (Figure 38). The mesostylid is very weak or absent with a relatively wide valley between the metaconid and the entoconid. This condition differs from species of Spermophilus such as S. shotwelli Black or S. argonautus Stirton and Goeriz (1942) that are anteroposteriorly compressed. The relative lack of anteroposterior compression resembles Black's (1963) description of S. wilsoni Shotwell. The entoconid is strongly connected to the posterolophid. The squirrel from the Warren Local Fauna tentatively is assigned to the genus Spermophilus. However, its specific identity is not clear. It is approximately the same size as S. gidleyi or S. argonautus and is smaller than S. wilsoni and S. shotwelli. It differs from both S. gidleyi and S. argonautus in having lower molars less anteroposteriorly compressed. This, as well as the incomplete metaloph, closed trigonid basin on M1, and open (?) trigonid on M2, most nearly resemble characters of Spermophilus wilsoni.
Family Geomyidae Bonaparte, 1845
Prothomomys new genus
Type Species: Prothomomys warrenensis new species. Etymology: Prefix Pro from Latin meaning before and thomomys from the existing name Thomomys Wied-Neuwied, 1837.
Prothomomys warrenensis n. sp. Diagnosis of Species: Same as for genus. Type series: UCMP 320186 – P4 is designated as the holotype. UCMP 320815 – DP4 and UCMP 320188 – M1 are designated as paratypes. Type locality: The holotype and both paratypes were collected from locality RV-7702.
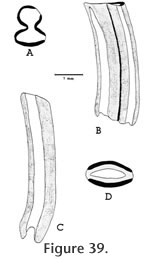
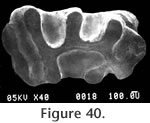
Localities: UCMP 320187 and UCMP 320595 - locality RV-6834, UCMP 123629 - locality V-6638, UCMP 124821 - locality V-2503. All other specimens are from locality RV-7702. Etymology: The species name is derived from warren indicating the Warren Local Fauna or Warren syncline and from –ensis denoting place. Description: Two permanent P4s are very hypsodont and rootless with well developed narrow dentine tracts. Both teeth are moderately worn with three of the four dentine tracts having breached the occlusal surface; the slightly shorter dentine tract is on the anterior lophid. The occlusal pattern of the P4 consists of a rounded anterior column narrowly connected to the antero-posteriorly compressed posterior column (Figure 39a and b). The anterior column will remain rounded throughout wear rather than becoming diamondshaped. The dP4 (UCMP 320185, Figure 40) is double rooted, low crowned, lophate, and has an occlusal pattern very similar to that of Parapliosaccomys oregonensis (Shotwell 1967). The molars of the Warren gopher are very hypsodont with well developed, narrow dentine tracts (Figure 39c and d). The molars are generally rootless although UCMP 320568 (M3) does have poorly developed roots and relatively shorter dentine tracts. The molars have enamel on both the anterior and posterior surfaces, and the anterior enamel plate is unreduced and continuous to the base of the tooth as in Thomomys. The occlusal shape of the molars is somewhat variable, but generally ellipsoid. UCMP 320564 (M1 or 2) has a marked lingual constriction. Discussion: The advanced degree of hypsodonty (primarily rootless) with presence of well developed narrow dentine tracts and continuous anterior enamel plates in both upper and lower molars suggest that Prothomomys n. gen. is closely related to, and probably ancestral to, the extant genus Thomomys Maximilian. Modern species of Thomomys are distinguished from other Geomyidae by having the lower molars with lingual constriction of lophids and upper molars with labial constriction of lophs. However, Zakrzewski (1969) noted that the majority of molars in Thomomys gidleyi, from the Blancan Hagerman Local Fauna, were not constricted, and Gustafson (1978) reported occlusal shape variable in molars of Thomomys sp. cf. T. gidleyi from the early Blancan White Bluffs Local Fauna. Prothomomys n. gen. differs from Parapliosaccomys oregonensis (Shotwell, 1967) whose dentinal tracts are irregular, the anterior enamel plate is reduced in upper and lower molars, and the anterior column of P4 becomes diamond-shaped with wear, rather than well rounded. Martin (1979) noted that "tear-shaped" molars are also developed in P. oregonensis from northern Oregon. The Warren gopher is very advanced with respect to the degree of hypsodonty and development of dentine tracts, and the dP4 suggests a close relationship with Parapliosaccomys oregonensis from the late Hemphillian of Oregon (Shotwell, 1967). Prothomomys n. gen. retains an isolated posterior lophid on DP4 that later becomes anteriorly connected in Thomomys. Various authors including Shotwell (1967), Zakrzewski (1969), and Martin (1979) have proposed that P. oregonensis was ancestral to Thomomys. If this were true, then the Warren gopher might represent an intermediate form between these two taxa. However, reduction of the anterior enamel plates on the lower molars of P. oregonensis makes it unlikely that P. oregonensis was ancestral to Thomomys. It is also unlikely that Prothomomys was derived from P. oregonensis because Prothomomys has continuous anterior enamel plates as in Thomomys. The Warren gopher therefore provides a potentially better ancestor for Thomomys than does P. oregonensis. The very close similarity in dP4s suggests that Prothomomys n. gen. was probably derived from Pliosaccomys as was Parapliosaccomys. These observations suggest a phylogeny in which both Prothomomys and Parapliosaccomys were derived from Pliosaccomys during the Hemphillian. Martin (1979) has convincingly demonstrated the evolution of Pliosaccomys through Parapliosaccomys in Oregon during the Hemphillian. This evolution was characterized by an increase in hypsodonty and the development of dentine tracts. In Parapliosaccomys oregonensis, however, the anterior enamel plates of the lower molars are strongly reduced. Prothomomys warrenensis also developed hypsodont cheek teeth with well defined dentine tracts and no roots. More importantly, P. warrenensis does not show reduction of the anterior enamel plates on the lower molars. Prothomomys may have been ancestral to Thomomys cf. T. gidleyi by the early Blancan (Ringold Formation, Gustafson, 1978). There are two described fossil species of Thomomys in North America: T. gidleyi Wilson (1933) (Hagerman, Idaho; San Timoteo Badlands, California; Ringold, Washington) and T. carsonensis Kelly (1994) (Fish Spring Flat Local Fauna, Nevada). All of these occurrences are in Blancan faunas. Unnamed species of Thomomys are reported from the Blancan faunas of the San Timoteo Badlands (Albright 1999), the Blancan Grand View Fauna, Idaho (Hearst 1998), the Blancan – Irvingtonian Anza-Borrego faunas of California (Becker and White 1981), the Late Hemphillian Santee and Devil's Nest Airport Local Faunas of Nebraska (Voorhies 1990), and the Clarendonian Iron Canyon and Dove Spring Faunas in California (Whistler and Burbank 1992). Tedford et al. (2004) list Thomomys (Plesiothomomys) as a taxon within the characterizing fauna of the late late Hemphillian (Hh4; Tedford et al. 2004). However, all described species of Plesiothomomys are from Pleistocene faunas. Suggested occurrences of Thomomys in the late Hemphillian and even Clarendonian may indicate a more complex phylogeny for Prothomomys, Thomomys, Pliosaccomys, and Parapliosaccomys, or an incomplete record of the ranges of these taxa.
Family Geomydiae Bonaparte, 1845
? Perognathus sp.
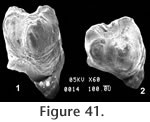
Locality: All specimens were collected from locality RV-7702. Description and Discussion: Three isolated teeth from the Warren Local Fauna are questionably assigned to the genus Perognathus Wied-Neuwied. The dimensions of the P4 (UCMP 320182) are square (0.83 mm x 0.83 mm) with the protolophid slightly narrower than the metalophid (terminology after Lindsay 1972). These lophids would unite medially with moderate wear. In early wear, the protolophid is v-shaped, however, the anteromedial groove shallows quickly. Both labial and lingual reentrants are well developed with the labial one slightly shallower. Other than size, the morphologies of UCMP 320184 (M1? 0.94 mm x 1.16 mm) and UCMP 320183 (M1 or 2: 0.94 mm x 1.01 mm) are of little value for identification. The P4 (UCMP 320182, Figure 41) is comparable in size and morphology to Perognathus gidleyi Hibbard from the early Blancan Rexroad Fauna, Kansas. It differs from the holotype of P. gidleyi in that the Warren specimen has no posteromedial groove, and the labial reentrant is slightly shallower than the lingual. UCMP 320182 differs from P. dunklei Hibbard by its larger size, from P. henryredfieldi Jacobs by its larger size and medial junction of the lophids, from P. mclaughlini Hibbard by its smaller size and medial junction of the lophids.
Family Muridae Illiger, 1811 Repomys gustelyi May, 1981 Repomys gustleyi is the most common rodent in the Warren Local Fauna (May 1981b). Repomys is a hypsodont cricetine that was originally described from three faunas in California and Nevada, and a complete description of Repomys from the Warren Local Fauna can be found in May (1981b). The etymology of this genus honors the late C.A. Repenning – i.e., Rep's mouse. The entire type series for Repomys gustleyi was collected from locality RV-7702 in Member Two of the Horned Toad Formation. May (1981b) identified two additional species of Repomys, R. panacaensis from the early Blancan Panaca Local Fauna in Nevada and R. maxumi from the early Blancan Maxum Local Fauna in California. Repomys has since been recognized at a number of additional localities in the southwestern USA including R. gustelyi from Devil's Nest Airstrip and Santee faunas in Nebraska (Voorhies 1990), R. gustleyi from the Mt. Eden Local Fauna (Albright 1999), R. maxumi from the San Timoteo Badlands (May and Repenning 1982; Albright 1999), R. panacaensis from the Buckhorn Local Fauna in New Mexico (Morgan and Lucas 2003), Repomys n. sp. from the Panaca Local Fauna (Lindsay et al. 2002), and R. arizonensis from Arizona (Tomida 1987). All known species of Repomys, except for R. gustleyi, are from Blancan age faunas. Most authors have followed May (1981b) in considering Repomys to have a North American origin (e.g., Jacobs and Lindsay 1984), although Voorhies (1990) lists Repomys as a latest Hemphillian immigrant with no supporting discussion. Tedford et al. (2004) listed Repomys (R. gustelyi) as one of the defining taxa for the late late Hemphillian (Hh4) based on the known occurrences in the Warren, Mt. Eden, Santee and Devil's Nest Airstrip Local Faunas. Genus POSTCOPEMYS Lindsay and Czaplewski (2011) Korth (1998) described the genus Antecalomys as a sigmodontine cricetid, interpreted phyletically between the genus Abelmoschomys from Clarendonian deposits in Florida described by Baskin (1986) and the extant cricetid genus Calomys (interpreted by Baskin to include the extinct taxon Bensonomys as a subgenus) recorded from numerous Blancan faunas in North America, and possibly part of the sigmodontine cricetid radiation that occurred in South America during the late Pliocene (but see Pardiñas et al. 2002). Part of the rationale for the genus Antecalomys is development of an oblique ridge in lower cheek teeth formed by alignment of the entolophulid with the posterior arm of the protocone (which are not aligned in Copemys), and retaining nonalignment of paraconule II with the anterior arm of the hypocone in upper cheek teeth, resulting in the absence of an oblique ridge in those teeth. Korth (1998) also noted a weakly divided anteroconid and presence of an accessory rootlet in M1 on the genotype of Antecalomys, A. phthanus. Accessory rootlets on upper and lower cheek teeth are characteristic of many sigmodontine rodents. Korth (1998) assigned two other known species, Copemys valensis (Shotwell 1967) and ?Copemys vasquezi (Jacobs 1977), to the genus Antecalomys, based primarily on development of an oblique ridge in lower teeth formed by alignment of the entolophulid with the posterior arm of the protocone and with nonalignment of paralophule II and the anterior arm of the hypocone in upper teeth. Lindsay and Czaplewski (2011) removed C. valensis and ?C vasquezi from the genus Antecalomys, considering that these taxa lack the development of accessory rootlets that are seen in some Sigmodontinae. They created the genus Postcopemys for the new species Postcopemys repenningi, from the early Blancan (and possibly late Hemphillian) Verde Formation in Arizona, and placed both C. valensis and ?C. vasquezi in the genus Postcopemys. Both Antecalomys and Postcopemys develop an oblique ridge in lower cheek teeth by alignment of the entolophulid and the posterior arm of the protocone, and lack an oblique ridge in upper cheek teeth (as in the genus Copemys) because protolophule II and the anterior arm of the hypocone are not aligned.
Postcopemys valensis
Shotwell (1967)
Locality: All specimens are from locality RV-7702. Description and Discussion: A small species of Postcopemys is represented by three brachydont cheek teeth (two M1s and one M1). UCMP 320576 and UCMP 320577 are both abraded anteriorly, but size and other dental features compare favorably to casts of Postcopemys valensis from Hemphillian deposits in Oregon. UCMP 320575 is unworn and of similar size. On the unworn specimen the anteroconid is very broad, gently rounded anteriorly (lacking a groove) and steep posteriorly where an anterolophid is directed posteriorly to join the metalophid on the midline. The metaconid is placed close to the lingual side of the anteroconid, partly closing the anterolingual sinus after moderate wear. Anterior arms of the protoconid and hypoconid are short, joining the metalophid I and the entolophid I, respectively. Posterior arms of the protoconid and hypoconid are long, with the protoconid arm joining the entolophid and the hypoconid arm expanding to become the prominent posterior cingulum that is flexed anteriorly to join the posterior base of the entoconid. The posterior arm of the protoconid and the entolophid are slightly offset, but with slight wear they become aligned, which is characteristic for both Antecalomys (Korth 1998) and Postcopemys (Lindsay and Czaplewski, 2011). A mesolophid and other accessory cusps and lophs are absent. Roots are not preserved on the M1.
Genus PEROMYSCUS Gloger, 1841
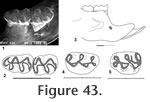
Locality: UCMP 126360 is from locality V-6638. All other specimens are from locality RV-7702. Description: A medium size, brachydont mouse with oblique cusp alignment in upper and lower molars is recorded from two mandibles, an isolated M1, and two M1 fragments. The occlusal outline of M1 is elongate oval, longer than wide, with four main cusps (protocone, paracone, hypocone, and metacone) and a large subequally bilobed anterocone. Labial cusps are placed opposite to the posterior side of lingual cusps. The anterior arm of the protocone is long, joining the posterior base of the anterocone centrally. The posterior arm of the protocone is short, joining the anterior side of protolophule II near the midline. The anterior arm of hypocone is long, joining the posterior side of protolophule II, with alignment of these lophs. The posterior arm of the hypoconid is long, directed labially, and joins the metalophule II to become a short posterior cingulum that terminates at the posterior side of the metacone. The protolophule II and metalophule II are short, as is a mesoloph that extends labially from the anterior arm of the hypocone, posterior to its union with protolophule II. A short lingual cingulum descends from the lingual lobe of the anterocone, terminating as a low cingulum that partly closes the anterolingual sinus. Three prominent roots, one lingual and two labial, are developed; accessory rootlets are unknown. The dentary includes a mental foramen that is relatively low and anterior, below the lowest position of the diastema, near the posterior ascension of the diastema. The masseteric crest terminates anteriorly below the anterior root of M1, well above the level of the mental foramen on UCMP 320570 and slightly above the level of the mental foramen on UCMP 320571. The occlusal outline of M1 is elongate triangular with a round anterior margin (Figure 43). Four main cusps (protoconid, metaconid, hypoconid, and entoconid) and a smaller, asymmetrical, single-cusped anteroconid are present. Lingual cusps are placed opposite to the anterior arms of labial cusps. The anterior arm of the protoconid is short, joining the posterior side of metalophid I that is directed anteriorly to join the posterior side of the anteroconid at the midline. The metaconid is placed relatively close to the anteroconid, and a narrow v-shaped valley separates these cusps. A long posterior arm of the protoconid joins the anterior side of entolophid I, and these lophs are aligned. A long anterior arm of the hypoconid joins the posterior side of entolophid I, while a long posterior arm of the hypoconid is flexed lingually to become a robust posterior cingulum that terminates short of the lingual tooth margin. There is no mesolophid, or other accessory styles or lophids. A labial anterior cingulum descends from the anteroconid, partly closing the anterolabial sinus; the labial sinus is partly closed by a low labial cingulum; lingual cingula are absent. Two well developed roots (anterior and posterior) are developed; accessory rootlets are unknown. The occlusal outline of M2 is a rounded rectangle, longer than wide, with four main cusps, as in M1 (Figure 43). Lingual cusps are placed opposite to the anterior arms of labial cusps, as in M1. The anterior arm of the protoconid is short, joining the posterior side of metalophid I that continues anteriorly to join the high anterior cingulum at the midline. The posterior arm of the protoconid is long; it joins the anterior side of entolophid I slightly lingual to the midline, and these lophids are aligned, as in M1. The anterior arm of the hypoconid is long; it joins the posterior side of entolophid I at the midline. The posterior arm of the hypoconid is long and flexed lingually to become a robust posterior cingulum that terminates short of the lingual tooth margin. There is no mesolophid or other accessory styles or lophids. The anterior labial cingulum descends gradually from the midline and is flexed posteriorly to terminate at the anterior base of the protoconid; there is no lingual anterior cingulum. The broad labial sinus is partly closed by a low labial cingulum; lingual cingula are absent. Two roots are developed as in M1; accessory rootlets are unknown. Peromyscus sp. A is most similar to P. kansasensis from the Rexroad Fauna in Kansas (Hibbard 1941). It differs from P. kansasensis by its slightly smaller size, however, a larger sample size may reveal overlapping size ranges. Peromyscus sp. A is larger that P. nosher from the White Bluffs Local Fauna, Washington; P. baumgarneri from the Fox Canyon and Rexroad Faunas, Kansas; P. cragini from the Cudahy Fauna, Kansas; and P. Hagermanensis from the Hagerman Local Fauna, Idaho; Deer Park Fauna, Kansas; and the 111 Ranch Fauna, Arizona. Peromyscus. sp A is smaller than P. antiquus from the Thousand Creek Fauna, Nevada, and P. pliocenicus from the Kern River Fauna, California. Miocene cricetid rodents in North America, commonly assigned to the genus Copemys, are characterized by non-alignment of oblique ridges on upper and lower cheek teeth. During the Pliocene cheek teeth of descendants of these Miocene cricetids gradually became aligned, primarily between the anterior arm of the hypocone and the paralophule II in upper teeth, and between the posterior arm of the protoconid and the entolophid I in lower teeth. The genus Antecalomys and the genus Postcopemys were both defined, in part, based on non-alignment of oblique lophs in upper teeth with alignment of oblique lophs in lower teeth. Most of the Pleistocene and extant species of Peromyscus can be characterized as having alignment of oblique lophs in both upper and lower teeth. In Antecalomys the upper teeth develop accessory rootlets, which are characteristic of some sigmodontine cricetids, and in Postcopemys none of the species develop accessory rootlets; Postcopemys is considered a probable member of the neotomine-peromyscine cricetid radiation, which also includes Copemys and the fossil and extant species of Peromyscus. The Warren Local Fauna is one of the few fossil records having species of both Postcopemys and Peromyscus as well as the hypsodont cricetid Repomys. Peromyscus sp. A may represent a new species of Peromyscus, however, due to the small sample size from the Warren Local Fauna, and the close comparison to P. kansasensis, we hesitate to name a new taxon for this material.
Peromyscus sp. B Material: UCMP 320572 – M1. Locality: UCMP 320572 is from RV-7702. Description and Discussion: A second species of Peromyscus is represented by an isolated M1 with clearly aligned anterior arm of the hypocone and paralophule II, as well as a bilobed anterocone. It is smaller than Peromyscus sp. A and the lobes of the anterocone are located closer together and separated by a shallower groove. The measurements of UCMP 320572 are: A.P. 1.71; T. 1.11.
Genus Jacobsomys Czaplewski, 1987
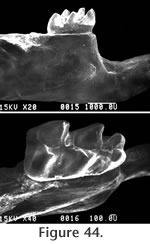
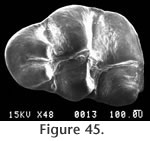
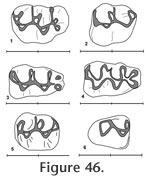
Figure 44,
Figure 45,
Figure 46
Paratype Material: UCMP 320579 – M3, UCMP 320580 – right maxillary fragment with M1, UCMP 320581 – left mandible fragment with M1, UCMP 320582 – M1, UCMP 320584 – M1, UCMP 320585 – M2, UCMP 320586 – M2, UCMP 320587 – M2, UCMP 320588 – M2, UCMP 320589 – M3, UCMP 320590 – partial M1, UCMP 320591 – partial M1, UCMP 320592 – partial M2, UCMP 320593 – M2. Referred Material: UCMP 320594 – M1. Type Locality: RV-7702. All specimens were collected from RV-7702 except for UCMP 320594 that was collected from RV-8103. Etymology: Named in honor of William R. Daily who collected and prepared most of the fossils in the original U.C. Riverside collection of the Warren Local Fauna.
M2 has a moderately well developed anterior cingulum and a strong posterior cingulum with a small mesostylid (Figure 46.5). A prominent mesolophid is present, and this tooth has two roots. The M1 has a strongly bilobed anterocone plus a long thin anteroloph and a mesoloph (Figure 45 and Figure 46). The primary cusps (protocone – paracone and hypocone-metacone) are arranged nearly opposite one another. The anterior mure joins the anterocone at the midline of the tooth. The mesoloph connects with the posterior side of the paracone forming a small enamel lake at the postero-medial corner of the paracone. A posterior cingulum is present but weak. At least three roots are present, but not well preserved on any of the specimens. The M2 has a strong anterior cingulum and a weak posterior cingulum (Figure 46.2). A mesoloph is directed labially from the mure along the anterior base of the paracone. The metalophulid II connects posteriorly with the hypocone. Czaplewski (1987) defined this genus on the basis of isolated teeth. The mandibular morphology for Jacobsomys dailyi can be described from the type specimen UCMP 320578 (Figure 44.1). The massateric crest is strong and terminates anteriorly beneath the anterior root of M1. The dorsal and ventral branches of this crest merge beneath the posterior edge of M1 and terminate at approximately the level of the mental foramen. The posterior edge of the diastema is very steep and the mental foramen is located dorsally. The depth of the mandible below M1 is 3.1 (+) mm. May (1981a) assigned this material to Calomys (Bensonomys) sp. cf. C. gidleyi. Following Czaplewski (1987), this material is better assigned to Jacobsomys and represents the earliest described species of this genus. Jacobsomys is further evidence of a rapid sigmodontine radiation in the late Miocene-early Pliocene in the southwestern USA. Repenning and May (1986) identified a sigmodont rodent from Blancan sediments near Truth or Consequences, New Mexico, as cf. Oryzomys. This material has since been referred to Jacobsomys sp. (Morgan and Lucas 2003), but the similarity with Oryzomys has led Lindsay and Czaplewski (2011) to suggest an ancestral relationship between Jacobsomys and Oryzomys. A partial M1 (UCMP 320594) is assigned to J. dailyi from locality RV-8103. The anterocone is missing, but the remainder of the tooth compares favorably with the M1's from locality RV-7702. Based on our interpretation of the magnetic polarity stratigraphy, RV-8103 is believed significantly younger than RV-7702 (Chron 3n.2r vs. Chron 3n.3r). J. dailyi may therefore extend from the latest Hemphillian into the earliest Blancan in the Horned Toad Formation pending eventual recognition of the mammal age boundary.
Order Carnivora Bowdich, 1821
Localities: UCMP 318700 - RV-8102. Description and Discussion: An associated right M1 (Figure 47.1) and a fragmentary left mandible (Figure 47.2) represent the borophagine dog Borophagus. Unfortunately, the diagnostic teeth of the mandible (P4 – M1) are fragmentary. Only the roots of P4 remain although it appears to have been inclined posteriorly and was a relatively large tooth with an anteroposterior length of approximately 17-18 mm. Only part of the labial side of M1 is preserved. The tooth was relatively large with an anteroposterior length of approximately 27.5 mm. M2 is missing, but a single alveolus is present. M3 is missing, and the alveolus is very small and slightly closed suggesting that the tooth was lost during the life of the animal. Judging from the poorly preserved alveoli, the premolars were strongly reduced. P1 is completely absent and P2-3 are represented by single, small alveoli. The alveolus for the canine is moderately large. The reduction of both M2-3 and especially the premolars anterior to P4 result in a short, stout mandible most of which was occupied by the fourth premolar, the first molar, and the canine. The tooth row is flexed labially. The upper dentition is represented by a single M1. The paracone is much larger than the metacone. A distinct anterolingual cingulum joins the anterior edge of the hypocone and projects anterior to the protoconule. The hypocone is well developed and is positioned directly transverse to the metacone. This specimen compares well with the M1 assigned to Osteoborus cf. cyonoides by Webb and Perrigo (1984) from the Hemphillian Corinto Fauna of El Salvador. This species has been allocated to Borophagus secundus by Wang et al. (1999), an assignment that is followed here. This material can certainly be assigned to the genus Borophagus. Wang et al. (1999: 296) indicate that B. secundus is the earliest species to show advanced features of Borophagus, such as a very short rostrum, broad palate, and a robust, high-crowned P4.
Family HEMIcyonidae Frick, 1926 Material: URC 18707 – edentulous fragment of right mandible, UCMP 29798 – upper premolar. Localities: UCMP 318707 - locality RV-8105 and UCMP 29798 - locality V-2503. Description and Discussion: A fragment of the right mandible of a large bear (UCMP 318707) contains the posterior alveolus of P3, both roots of P4, and the anterior alveolus of M1. UCMP 318707 compares well with USNM 264246, a nearly complete right mandible of Agriotherium sp. cf. A. silvalensis collected from the lower Coso Formation 90 miles north of the Horned Toad Hills. Richard Tedford (AMNH, personal commun., 1980) previously assigned a single rooted, bulbous upper premolar (UCMP 29798) from the Warren Local Fauna to the genus Agriotherium. Agriotherium was also reported by Frick (1921) from the late Hemphillian Mt. Eden beds of southern California. Agriotherium appears to have been the common bear in the southern USA and northern Mexico during the late Hemphillian (Hunt 1998), and no reports of Indarctos are known from this area. Tedford et al. (2004) include Agriotherium as an immigrant taxon that first appears in North America in the late Hemphillian (Hh3).
Family Felidae
Fischer de Waldheim, 1817 Material: UCMP 318701 – partial upper canine, UCMP 318702 – proximal phalanx, UCMP 318703 – 1st metacarpal, UCMP 318704 – 4th metacarpal, UCMP 320103 – distal metacarpal, UCMP 21222 – proximal phalanx. Localities: UCMP 318701 and UCMP 320103 – locality RV-7702; UCMP 318702 and 318704 – locality RV-8107; UCMP 318703 – locality RV-7504; UCMP 21222 – locality V-1750. Description and Discussion: A large machairodont cat is represented in the Warren Local Fauna by a partial upper canine and five foot-elements. Only the root portion of the upper canine was recovered and the greatest anteroposterior length of this tooth is 32.5 mm. This measurement is at the broken edge, so it is a minimum estimate. The size of this tooth compares well with measurements of canines assigned to Machairodus coloradensis Cook reported by Dalquest (1969) from the Coffee Ranch Local Fauna. The foot elements of the Warren cat are also referable to M. coloradensis as figured by Burt (1931). Martin and Schultz (1975) suggested that the holotype of Machairodus catacopis Cope was actually the pseudaelurine cat Nimravides. They recognized M. coloradensis as the only valid species of Machairodus in North America (see also Martin 1998). The large machairodont from the Coffee Ranch Quarry, commonly assigned to M. catacopis (Dalquest 1969), is now recognized as M. coloradensis. The machairodont cat from the Warren Local Fauna is therefore assigned to M. coloradensis, as also cited by Martin (1998), who recorded this taxon in late Hemphillian faunas from the Kansas-Texas panhandle region to Arizona and California. Machairodus is not known from North American faunas younger than late late Hemphillian (Hh4) (Tedford et al. 2004).
Order URANOTHERIA
McKenna and Bell, 1997
Material not assigned to this specimen includes UCMP 75438 – LM3, UCMP 316980 – dP3?, UCMP 316979 – dP2?, and LACM 121968 – deciduous lower tusk.
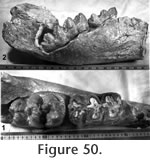
Description: The unequivocal association and excellent preservation of the elements of UCMP 313449 make this a very important specimen. The nearly complete mandible (Figure 48, Figure 49, Figure 50) is relatively short with the symphyseal portion downturned at an angle of approximately 50 degrees from the plane of the tooth row. The total length of the dorsal and symphyseal furrow is 28.0 cm with an average internal width of 52.0 mm. The depth of the mandible beneath the anterior alveolus of M1 is 14.5 cm and at the anterior edge of the ascending ramus is 11.1 cm. The greatest transverse width of the dentary is 13.0 cm just anterior to the ascending ramus. The maximum outside transverse dimension of the complete mandible near the base of the ascending rami is 34.5 cm.
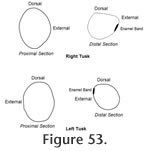
The symphyseal structure of the right dentary is complete with the proximal end of the right tusk in place. The maximum observable length of the right tusk is 44.0 cm. The tusk is transversely compressed near its alveolar margin with a dorso-ventral diameter of 55.5 mm and a transverse diameter of 43.8 mm. The cross-sectional shape of the tusk changes toward the tip with a distinct flattening of the dorsal surface (Figure 53). The ventral surface of the tusk is well worn for approximately 8.0 cm from the tip. A straight enamel band extends 18.8 cm posteriorly from the tip along the external side of the tusk. This band is only 14.0 mm at its widest point and tapers slightly posteriorly where it terminates in a feathered intergrowth with the external dentinal surface. The anterior portion of the enamel band is polished by wear, but otherwise the texture is rough.
The M2's are in full occlusion with dentine apparent on all three lophids (Figure 50.2). A simple trefoil pattern is preserved on the third labial hemilophid and was presumably present on the first and second at an earlier stage of wear. The crowns of the M3's are fully developed, but at the time of death, these teeth had not advanced forward into occlusion (Figure 50.150.2). Removal of bone from the internal side of the left dentary shows the posterior portion of LM3 (Figure 50.1). This tooth has four strong lophids and an incipient fifth lophid consisting of one major and two minor conulids. Simple trefoils are present on the four major labial hemilophids. An external cingulum is present extending from the anterolabial edge of the tooth back to the base of the second lophid, beyond which the tooth is hidden. The well preserved upper tusks are spiral with wide enamel bands extending their complete length. The right tusk is 130 cm long, and the maximum width of its enamel band is 71.5 mm. The enamel band follows the spiral of the tusk from the proximal-external side around the ventral surface to the distal-internal side. The total length of the left tusk is 138.5 cm, and the maximum width of its enamel band is 78.6 mm. The left tusk spirals in the same sense, although symmetrically opposed to the right tusk. Transverse compression at the proximal end follows the spiral of the upper tusks, but the cross-section becomes more circular distally. The maximum dimensions near the middle of the right tusk are 10.3 cm x 7.9 cm. Both tusks are slightly curved so that in a reconstructed orientation they would be curved outward in top view and concave down in side view. The enamel band of the upper tusks has a rough, pebbly texture as in the lowers, but is worn nearly smooth along the anterior third of the tusk.
The two M3's of UCMP 313449 are completely unworn, but with wear, would exhibit simple trefoils on all four lingual hemilophs. The second labial hemiloph would also develop a trefoil pattern with moderate wear. The trefoil pattern of the fourth internal hemiloph of UCMP 75438 is less well developed than in UCMP 313449. Both specimens exhibit a small rugose heel projecting labially from the posterior extension of the fourth lingual hemiloph. LACM 129168 is a very small and presumably deciduous lower tusk. The tusk is transversely compressed and curves upward (?). Its slight lateral curvature suggests that it was from the right dentary and, therefore, the external surface is best preserved. There is no trace of an enamel band. Numerous longitudinal ridges extend down the tusk resembling the "corduroy" texture described by Olsen (1957). The post-cranial material of UCMP 313449 is not described. These specimens are mounted in the Geology Department at U.C. Riverside. Discussion: May (1981a) originally assigned the gomphotheriid mastodont from the Warren Local Fauna to the genus Cuvieronius, based on his assessment that Rhynchotherium was nomen dubium. Nearly every author who has described Rhynchotherium and/or Cuvieronius fossils argues that the taxonomic history of these genera is both confusing and complicated (Miller 1980; Webb and Perrigo 1984; Miller 1990; Lambert 1996; Corona-M and Alberdi 2006). However, most authors continue to use these generic names and Lambert and Shoshani (1998) diagnose Rhynchotherium as having variably spiral upper tusks with enamel bands; lower tusks that are medio-laterally compressed and commonly have enamel bands; and mandibular symphysis longirostrine, but significantly deflected downward. They characterize Cuvieronius as brevirostrine and lacking lower tusks, but sharing the characters of spiral upper tusks with enamel bands and molar morphology with Rhynchotherium. The gomphotheriid material from the Warren Local Fauna is assignable to the genus Rhynchotherium on the basis of its spiral upper tusks with enamel bands, relatively simple molars, mandible with downturned symphysis, and laterally compressed lower tusks with external enamel bands. The material compares reasonably well with R. edensis from Mt. Eden (Frick 1933). However, the enamel bands on the lower tusks are quite reduced with respect to R. edensis. The Warren material is therefore referred to as Rhynchotherium sp. cf. R. edensis. The molar morphology of these gomphotheriid taxa exhibit large variability, and specific characteristics are unclear. R. sp. cf. R. edensis does possess a pentalophid on M3 consisting of three conulids like R. edensis. R. falconeri reportedly has two conulids in this position, and R. browni only has a single conulid. The significance of this variability is not clear. The upper tusks of UCMP 313449 are spiral with a wide enamel band. Miller (1980) also reports a similar upper tusk from the early Blancan, Las Tunas fauna of Baja, California. The only completely observable M3 of R. sp. cf. R. edensis has three conulids forming the posterior heel and a length-width index of 43. Both of these parameters compare well with topotypic R. edensis. Webb and Tessman (1968) and Miller (1980) use these criteria to separate species, but it remains unclear whether these characters are diagnostic of different species. The lower tusks of UCMP 313449 are laterally compressed as in R. edensis, but the external enamel band on these tusks is much narrower and shorter. Believing the Warren Local Fauna to be older than Mt. Eden, May and Repenning (1982) suggested that the reduced enamel band on the Warren Rhynchotherium (their Cuvieronius), represented a more primitive condition. Webb and Perrigo (1984) disagreed with this hypothesis as material assigned to R. blicki from the late Clarendonian to early Hemphillian Gracias Fauna of Honduras possess laterally compressed lower tusks with a broad enamel band. Furthermore, these authors disagree with the assignment of the Mt. Eden gomphotheriid to Cuvieronius, as they believe this genus is characterized by the lack of lower tusks. They suggest that the New World gomphotheres evolved from longirostrines by deflection of the symphysis, then by shortening and thickening the mandible and finally by losing the lower tusks. The reduced enamel band in R. sp. cf. R. edensis from Warren might therefore be more derived than R. edensis from Mt. Eden, however the neotype of the latter figured by Frick (1933) appears to be more brevirostrine with a symphyseal angle of approximately 60° rather than the 50° observed in the Warren form. The symphyseal angle of the more primitive R. blicki from the Gracias Fauna is reported by Webb and Perrigo (1984) as approximately 25°, however, based on their figure 4, the angle measured relative to the tooth row appears to be closer to 40°. Rhynchotherium sp. cf. R. edensis also compares favorably with R. cf. R. falconeri described by Miller (1980) from the early Blancan Las Tunas Local Fauna (Baja, California). This form has a continuous enamel band on the lower tusks that is narrower than in R. edensis (20 mm vs. 31 mm), but has a less abbreviated mandible and a symphyseal angle of 56°. As described above, the enamel band on the lower tusks of R. sp. cf. R. edensis is 7-14 mm wide and only present on the distal half of the tusk. Miller (1980) suggested that R. edensis, R. falconeri, and R. browni may all represent a single species, but he did not formally propose synonymy. Miller (1990) described a skull and mandible of Rhynchotherium from the Gila River area of southeastern Arizona that he believes is probably latest Pliocene in age. Unfortunately, the specimen was collected without locality information so the stratigraphic provenance and age is uncertain. Nevertheless, this specimen possesses lower tusks that lack enamel bands in a brevirostrine mandible with a strongly deflected symphysis (65°). Unlike older specimens of Rhyncotherium that possess enamel bands on the lower tusks, the upper and lower tusks of the Gila River Rhynchotherium never came into occlusion. Miller (1990) suggested a probable phylogeny progressing from Gomphotherium to Rhynchotherium to Cuvieronius.
Family Mammutidae Hay, 1922
Localities: UCMP 318112 – locality RV 8126, UCMP 318113 – locality RV 7148, UCMP 318114 – locality RV 8112, UCMP 318115 – locality RV 8128, UCMP 320613 – locality RV 8114, UCMP 320614 – locality RV 7702. Description and Discussion: A second proboscidean is represented in the Warren Local Fauna by six specimens, most of which are teeth. A moderately well worn, left M2 (UCMP 318115) is trilophid with clean valleys and lacks accessory conulids on the lophids. Anterior and posterior cingula are present, but these are small and without accessory conulids. The tooth widens posteriorly.
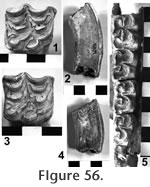
In contrast, the M3 of UCMP 38115 (Figure 55) has a posterior heel developed as a narrow ridge with five to six small conulids. This heel is also much closer to the fourth major lophid causing the tooth to appear relatively wider than UCMP 318112. This qualitative appearance is quantitatively expressed as the greater width-length index of 53. The valleys lack accessory conulids and are partially filled with cement. A moderately worn right P4 (UCMP 318114) is bilophid with strong anterior and posterior cingula that are ornamented with small accessory conulids. The anterior portion of a left mandible is preserved on UCMP 318115 with no suggestion of an alveolus for a lower tusk. Madden (1977, 1980) has presented brief reviews of Pliomastodon concluding that there are only two valid species in North America, namely P. matthewi and P. vexillarius. Madden (1980) states "... (late Hemphillian to late Blancan) P. vexillarius is distinguished from progenitor P. matthewi by its rounded upper tusks, tuskless mandibles, and 4-5 ridged M3." The material from the Warren Local Fauna is consistent with this diagnosis of P. vexillarius. The M3's of UCMP 318112 are quite advanced with a well developed pentalophid and a low width-length index. The seeming disparity between these M3's and UCMP 318115 is attributed to either sexual dimorphism or a large normal interspecific variation. Lambert and Shoshani (1998) consider Pliomastodon Osborn (1926) to be a junior synonym of Mammut Blumenbach (1799), and they include only two valid species: M. americanum and M. matthewi. Mammut vexillarius (misspelled vexillanus on p. 610 of Lambert and Shoshani 1998) is assigned to Mammut sp. along with all other named species. These authors suggest that Mammut extends from the Clarendonian to the terminal Pleistocene and was uncommon yet widespread across North America during the Clarendonian and Hemphillian "where it is commonly referred to as Pliomastodon in the literature." We have followed this practice, using the name Pliomastodon.
Order Perissodactyla Owen, 1848
Localities: UCMP 29778 – locality V2503, UCMP 29785 – locality V-2503, UCMP 320010 – RV-8125, UCMP 320050 – locality RV-8110, UCMP 320011 – locality RV-8112, UCMP 320052 – locality RV-7702, UCMP 320057 – locality RV-8113, UCMP 320058 – locality RV-8114, UCMP 320054 – locality RV-7148, UCMP 320048 – locality RV-8115, UCMP 320016 – locality RV-8115, UCMP 320015 – locality RV-8107, UCMP 320019 – locality RV-8105, UCMP 320017 – locality RV-8110, UCMP 29787 – locality V-2503, UCMP 320014 – locality RV-8117, UCMP 320018 - RV-8107, UCMP 320049 – locality RV-8125, UCMP 320013 – locality RV-6834, UCMP 320012 – locality RV-8118, UCMP 320051 – locality RV-8120, UCMP 320053 – locality RV-7702, UCMP 320055 – locality RV-8114, UCMP 320056 – locality RV-8105, UCMP 29775 – locality V-2503, UCMP 320060 – locality RV-8126, UCMP 320061 – locality RV-8126, UCMP 29784 – locality RV-2503, UCMP 320043 – locality RV-6834, UCMP 320046 – locality RV-7702, UCMP 320038 – locality RV-7702, UCMP 320035 – locality RV-8114, UCMP 320044 – locality RV-8107, UCMP 320040 – locality RV-8110, UCMP 320031 – locality RV-6834, UCMP 320598 – locality RV-7702, UCMP 320022 – locality RV-8127, UCMP 320033 – locality RV-8105, UCMP 320030 – locality RV-8112, UCMP 320039 – locality RV-8110, UCMP 320024 – locality RV-7702, UCMP 320036 – locality RV-8107, UCMP 320032 – locality RV-7702, UCMP 320023 – locality RV-8107, UCMP 29781 – locality V-2503, UCMP 320026 – locality RV-8114, UCMP 320027 – locality RV-8121, UCMP 320037 – locality RV-8110, UCMP 320042 – locality RV-8110, UCMP 320020 – locality RV-7148, UCMP 29775 – locality V-2503, UCMP 320028 – locality RV-8107, UCMP 320029 – locality RV-8107, UCMP 320047 – locality RV-7702, UCMP 320041 – locality RV-8105, UCMP 123628 – locality V-6638, UCMP 320059 – locality RV-8105, UCMP 320021 – locality RV-8126, UCMP 320063 – locality RV-8126, UCMP 320064 – locality RV-8126, UCMP 320065 – locality RV-8126, UCMP 320066 – locality RV-8126, UCMP 320067 – locality RV-8105, UCMP 320068 – locality RV-8125, UCMP 320069 – locality RV-8122, UCMP 320070 – locality RV-8107, UCMP 320071 – locality RV-7702, UCMP 320073 – locality RV-8123, UCMP 320076 – locality RV-8110, UCMP 320077 – locality RV-7702, UCMP 320078 – locality RV-8124, UCMP 320079 – locality RV-8105, UCMP 320080 – locality RV-8120, UCMP 320081 – locality RV-7702, UCMP 320082 – locality RV-M799, UCMP 320083 – locality RV-7702, UCMP 32084 – locality RV-8120, UCMP 320085 – locality RV-7702, UCMP 320086 – locality RV-7702. Description and Discussion: A single horse from the Warren Local Fauna is represented predominantly by isolated teeth and post-cranial material. Four partial mandibles contain associated lower teeth. However, no cranial specimens have been recovered. This material compares favorably with Dinohippus edensis Frick from the late Hemphillian Mt. Eden Local Fauna in California. Measurements of the cheek teeth are shown in Table 10. The Warren horse is similar in size to Dinohippus edensis, D. osborni, D. mexicanus and D. interpolatus. The upper cheek teeth are slightly to moderately curved, similar to D. mexicanus or D. osborni, but less than D. interpolatus (Figure 56). The protocone is generally oval in shape with no lingual groove and no anterior projection. The pre-protoconal groove is shallow and projects externally toward the area between the mesostyle and metastyle. These characters of the protocone are considered primitive and distinguish the Warren horse from both D. mexicanus and D. osborni. Large samples of D. interpolatus exhibit variation in protocone shape from the primitive condition similar to D. edensis, to some with an incipient anterior projection. The fossette borders of the upper molars exhibit simple plications including a pli-protoloph which become obliterated with extended wear. A pli-caballin is present on the premolars but absent on the molars, as generally common in Neogene equids. The styles are moderately developed as in D. edensis but smaller than D. mexicanus. The lower premolars are considerably larger than the molars (Figure 56.5). The metaconid is slightly larger than the metastylid although more nearly equal than in the molars. The metastylid of P2 is much larger than the metaconid. The entoconid is relatively large and round to oval in shape. The entoflexid is well developed with strong anterior and posterior projections while the metaflexid has only a slight anterior projection on P2 and only posterior projections on P3 and P4. The labial walls of the protoconid and hypoconid are rounded as in D. edensis rather than flattened as in D. mexicanus. In the lower molars, the metaconid is much larger than the metastylid with only a very shallow lingual valley separating these cusps. The labial reentrant projects lingually between the entoflexid and the metaflexid. A pli-caballinid is never present. In two of the six M3's, the posterior enamel loop is completely isolated as a lake (Figure 53.5), a condition that does not appear to be a result of wear as other specimens (at comparable wear stages) show a joined posterior loop. The horse from the Warren Local Fauna is similar in size to D. edensis, D. osborni, D. mexicanus, and D. interpolatus. It differs from D. mexicanus in having a more primitive protocone, less complex fossette borders, weaker styles, rounded labial walls on the protoconids and hypoconids and a better developed linguaflexid (MacFadden 1984). It differs from D. interpolatus in having less curved upper cheek teeth and in never exhibiting an advanced protocone. It differs from D. osborni in having a more primitive protocone. Lance (1950) suggested that the variation seen in the large sample of D. mexicanus from the Yepom,era was as great as the differences described by Frick (1921) in separating D. osborni from D. edensis. A major distinction between these two species is the stage of development of the protocone. MacFadden (1998) recognized both D. osborni and D. edensis as valid species of Dinohippus. The horse from the Warren Local Fauna resembles D. edensis very favorably but never exhibits the derived protocone of D. osborni.
Family Rhinocerotidae Gray, 1821
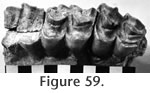
Localities: UCMP 318705 - locality RV-8114, UCMP 316981 - locality RV-7702, UCMP 29797 - locality V-2503. Description and Discussion: Three specimens from the Warren Local Fauna are identified as the rhino Teleoceras Hatcher. The most diagnostic of these is a well worn, partially fragmented, left P2 (Figure 57.3). It compares favorably with Teleoceras fossiger from Long Island, Kansas, and with the T. fossiger skull figured by Matthew (1932) from Brown County, Nebraska. It is also similar to the P2 of T. cf. fossiger figured by Stirton (1939) from the Pinole Tuff Local Fauna, California. The M1? is extremely worn although it is consistent with the size of T. fossiger. A single metapodial is referable to Teleoceras (Figure 54.1 and 54.2). The fragments of rhino material from the Warren Local Fauna preclude more detailed discussion of specific relationships: it appears most similar to T. fossiger from Hemphillian deposits in California and the Great Plains. Tanner (1975) has emphasized the evolutionary increase in size from the Valentinian Teleoceras sp. (Skinner et al., 1968) to T. schultzi Tanner from the late Hemphillian. However, Prothero (2005) suggested that T. fossiger is restricted to the early Hemphillian, and late Hemphillian forms of this genus are smaller. The few specimens from the Warren Local Fauna are within the known size range of Hemphillian forms.
Order Artiodactyla Owen, 1848 Material: UCMP 318706 – upper canine, UCMP 29799 – astragalus Localities: UCMP 318706 – locality RV-6834, UCMP 29799 - locality V-2503. Description and Discussion: The peccary Prosthennops Gidley is questionably represented by an upper canine and an astragalus. The canine compares favorably with Prosthennops kernensis from the Kern River Formation of California. Wright (1998) lists Prosthennops kernensis among a group of indeterminate Neogene tayassuids.
Family Camelidae Gray, 1821
Localities: UCMP 320090 – locality RV-8110, UCMP 320091 – locality RV-6834, UCMP 320092 – locality RV-8107, UCMP 320093 – locality RV-8129, UCMP 320094 – locality RV-8107, UCMP 320095 – locality RV-8112, UCMP 320096 – locality RV-7702, UCMP 320098 – locality RV-8107, UCMP 29788 – locality V2503, UCMP 29790 – locality V-2503, LACM 1552 – locality LACM 1552.
Entostyles are absent on all three upper molars as well as on UCMP 320095 (M2), although small cingula are present on both sides of the valley between the protocone and the metaconule near the base of the enamel. Both M3's lack anterior stylids. Harrison (1985) grouped the large camels Megatylopus and Titanotylopus together in the Tribe Camelini united primarily by a series of mandibular and cranial features. Very little of the material from the Warren Local Fauna provides diagnostic evidence for separation of these two taxa. Titanotylopus is generally considered to have less hypsodont cheek teeth and larger body size than Megatylopus. In his review of Megatylopus, Webb (1965) recognized four species that are diagnosed primarily on premolar characters and cranial morphology. However, Harrison (1985) recognized only two valid species (M. gigas and M. matthewi) distinguished by P3 morphology and the depth of the maxillary fossa. The holotype of Megatylopus matthewi from the late Hemphillian Coffee Ranch Local Fauna compares favorably with UCMP 320094. This comparison includes overall size, as well as dental morphology. Webb (1965) states that the upper molars are of little diagnostic value, however, he describes M. matthewi as having well developed parastyles, mesostyles, and anterolabial ribs, all of which are observed in the Warren Megatylopus. Megatylopus ranges from the late Clarendonian to the early Blancan in the western USA and M. matthewi has been described from the Hemphillian Wikieup Local Fauna, Arizona, the Chamita Formation, New Mexico, and the El Ocote Local Fauna, Mexico.
Genus Hemiauchenia
Gervais and Ameghino, 1880
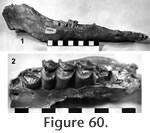
Localities: UCMP 320612, and UCMP 320131 – locality RV-8129; UCMP 320611 and UCMP 320117 – locality RV-6834; UCMP 320126 and UCMP 320121 – locality RV-7702; UCMP 320124 – locality RV-8112; UCMP 320127 – locality RV-8114; UCMP 29794 and UCMP 29791 – locality V-2503. Description and Discussion: A medium size, lamine camel in the Warren Local Fauna is identified as Hemiauchenia vera Matthew. The best specimen, UCMP 320612 (Figure 60), includes associated left and right mandibles and the left maxillary of an advanced juvenile with P3 just erupting, C1 in the process of being replaced, and three alveoli of dP4 still present. The right mandible is nearly complete except for fragmentary dentition. The anterior portion of the left mandible is missing. UCMP 29794 is a fragmentary right mandible with P4-M2 of a mature adult. The lower dental formula of this taxon is I1-3, C1, P1, 3, 4, and M1-3. The lower molars are relatively low crowned, reflected in the shallow depth of the mandible. Diagnostic characters of the lower dentition include the small canine, the lack of a P2, the length of the P1-3 diastema (40 mm), the morphology of P4, and the moderately strong anterior stylids ("llama buttresses") on M2 and M3. The P4 of UCMP 29794 is angular with a single posterior fossetid and a strong anterolingual groove. These characters, along with the sharp diastemal crest and the long slender metapodial (UCMP 320611), permit confident assignment to the genus Hemiauchenia. The P4-M3 length is difficult to assess accurately because the M3 is absent on UCMP 29794, and the permanent P4 is absent on UCMP 320612. The size of the Warren Hemiauchenia appears to be slightly larger than the holotype of H. vera, although measurements of individual teeth (e.g., M2) are very similar. The upper dentition is best represented by a maxillary (UCMP 320612) with P4-M3 (Figure 60.2). The permanent P4 has erupted but is essentially unworn, as is the M3. Parastyles and mesostyles are relatively strong with moderately developed external ribs. Entostyles are absent. Breyer (1977) reviewed the genus Hemiauchenia and recognized three North American species including H. vera, H. blancoensis, and H. macrophela. Breyer concluded that these three species can be separated based on the post-P1 diastemal length, with post-P1 diastemal length of 35 mm in the holotype of H. vera, 50 mm in H. blancoensis, and 60 mm in H. macrophela (Breyer 1977). The latter two species are known from Blancan and Pleistocene faunas of North America, respectively. Post-P1 diastemal length in the Warren Hemiauchenia is 40 mm, placing it in H. vera. Honey et al. (1998) continued to recognize these three species and follow the suggestion of Webb (1974) that Tanupolama is a junior synonym of Hemiauchenia, and Webb et al. (1981) that "H." minima from Florida (Mixon Bone Bed and Love Local Faunas) deserves a new generic name. The primitive hypsodonty, the shallowness of the mandible, the moderate development of the anterior stylids on M2-3, the short post-P1 diastema, and the retention of P3 allow taxonomic assignment of the Warren lamine to Hemiauchenia vera. H. vera is known from Hemphillian faunas of North America including the Ogallala Group near Long Island, Kansas; Coffee Ranch Local Fauna, Texas; Bone Valley, Florida; San Juan – Rak Camel Quarries Horizon of the Chamita Formation, New Mexico; Wikieup Local Fauna, Arizona; Yerrington, Nevada and the Modesto Reservoir Local Fauna, California.
CAMELIDAE
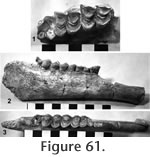
Localities: UCMP 320110 and UCMP 320089 – locality RV-8129, UCMP 320115 and UCMP 320118 – locality RV-8114, UCMP 320113 and UCMP 320123 – locality RV-6834, UCMP 320088 and UCMP 320120 – locality RV-7148, UCMP 320114 and UCMP 320127 – locality RV-7702, UCMP 320119 – locality RV-8121, UCMP 29795 – locality V-2503. Description and Discussion: A third camelid of uncertain identity is present in the Warren Local Fauna. UCMP 320110 (Figure 61) is a nearly complete right mandible extending from the posterior edge of the symphysis to the base of the ascending ramus with P4-M3 and no alveoli for P2 or P3. The diastemal crest is not as sharp as in Hemiauchenia vera. The absence of P3 is an advanced character observed in Camelops, Lama, and Camelus although Webb (1974) notes that the presence of P3 is variable in Pleistocene Hemiauchenia. The teeth are well worn, and no fossetid is observable on P4. This tooth differs from that of the larger Hemiauchenia vera by being less angular with a weaker anterolingual groove. The M1 is worn nearly to the base of the enamel and is of little diagnostic value. Anterior stylids are absent on all lower molars of these specimens except for UCMP 320115, a lower M3 with a weak anterolingual stylid that is referred to this taxon on the basis of size. The upper molars are undiagnostic except for their relatively low height of crown (Figure 61.1). Following Harrison (1979), only one genus belonging to the subfamily Camelini has P2 and P3 absent and also lacks anterior stylids on the lower molars: the Pleistocene to recent Eurasian genus, Camelus. Webb (1965) suggested that "some large, late Clarendonian species of Procamelus, like P. grandis or P. occidentalis is the most likely North American ancestor to the extant Asian camels" (i.e., Camelus). No relationship is suggested between the small Warren camelid and Camelus. The small Warren camelid differs from Procamelus by lacking P2-3. In Lama, the P2 and P3 are lost, but the anteroexternal stylids on the lower molars are very strong. In summary, the affinities of this small Warren camelid are uncertain, however, it appears to be morphologically advanced relative to known Hemphillian forms.
Family Antilocapridae Gray, 1866 Material: UCMP 29800 – miscellaneous postcranial elements including an astragalus, proximal phalanx, and distal metapodial fragments; UCMP 29801 – two upper premolars. Locality: UCMP 29800 and 29801 – locality V-2503. Description and Discussion: Lacking horn cores, it is extremely difficult to identify late Tertiary antilocaprids even at the generic level. An uncertain identification of cf. Sphenophalos is made in this case because the Warren material is indistinguishable from Sphenophalos specimens from Thousand Creek, Nevada, that were examined in the collections at the Los Angeles County Museum of Natural History. Biostratigraphy
May (1981a) and earlier references considered all of the fossils from the Horned Toad Formation as the Warren Local Fauna. Based on the subsequent recognition of the Lawlor Tuff superjacent to most of the fossils in the Warren Local Fauna, with an age close to the Hemphillian/Blancan boundary, it is important to analyze the fossils from above this stratigraphic marker for evolutionary differences. At present, only two taxa are identifiable from localities above the Lawlor Tuff including Jacobsomys dailyi and Dinohippus edensis. Specimens of these taxa are limited and do not provide evidence for a more advanced stage of evolution than those below the Lawlor Tuff. Guided by M. Woodburne, B. Reynolds and volunteers from the Mojave River Valley Museum in Barstow, California, collected a large volume of matrix from a locality along the north limb of the Warren syncline in the lower part of Member Four above the Lawlor Tuff. Numerous bone fragments and a limb bone of a small mammal were found during surface prospecting at this locality in 2006, although no identifiable fossils have yet been recovered from the washed matrix. The majority of fossil mammal localities in Member Two are from sediments exhibiting reversed magnetic polarity (Figure 62). Sites RV-8105 and RV-8120 are shown correlative with the lower part of the overlying normal polarity magnetozone. However, no paleomagnetic samples were collected at these localities, and the lowest definitive normal polarity site is the Lawlor Tuff that occurs stratigraphically above these localities. The suggested correlation of these fossil localities with the lower part of Chron 3n.3n is simply based on the arbitrary placement of the polarity boundary midway between the highest reversed polarity site and the lowest normal polarity site. Taxa from these sites include Dinohippus edensis that is also found above the Lawlor Tuff and cf. Agriotherium. A single locality (RV-8131) was discovered in the lower strata of Member One and may record a fauna distinct from the Warren Local Fauna. This suggestion is based on a single fragment of an upper cheek tooth of a horse that has very complex fossette borders and is unlike the teeth of Dinohippus edensis. Age and CorrelationTwenty-four mammalian taxa have been identified in the Warren Local Fauna. A late Hemphillian age is based on comparison with other faunas and assessment of the stage of evolution of the fossils. The occurrence of Cryptotis sp. in the Warren Local Fauna represents a range extension of this genus. Lindsay et al. (1984) pointed out that shrews are rare in Hemphillian faunas, becoming more common and diverse in the Blancan. Repenning (1967) incorrectly assigned a shrew from the late Hemphillian Christmas Valley Local Fauna of Oregon to Cryptotis adamsi. The oldest irrefutable occurrence of C. adamsi would be from the Blancan Rexroad Fauna of Kansas (Hibbard 1953). Wagner (1981) reported Cryptotis from the late Hemphillian Modesto Reservoir Local Fauna. However, no articular condyle or coronoid process is known from these specimens (Wagner, personal commun., 1980). As discussed in the systematics section, Cryptotis sp. from Warren shows a number of characters that are interpreted as being more primitive than C. adamsi. The material assigned to cf. Scapanus is of no help in determining the age of the fauna. Scapanus ranges from Clarendonian to Recent faunas in North America. Hypolagus vetus occurs in both early and late Hemphillian faunas of North America. White (1987) considers H. vetus to be restricted to the Hemphillian and Blancan of the western USA and Baja, California. Hypolagus edensis is known from the late Hemphillian, Redington and Mt. Eden Local Faunas (Arizona and California), as well as from a number of Local Faunas of Blancan age including Hagerman (Idaho) and Taunton (Washington) (White 1987). The new geomyid Prothomomys warrenensis is very advanced relative to most Hemphillian gophers. In development of hypsodonty, P. warrenensis approaches the morphology of the Blancan to Recent genus Thomomys. Prothomomys provides a potential ancestor for Thomomys gidleyi, the oldest record of which is early Blancan (Gustafson 1978). A marked similarity in dP4 morphology suggests a close relationship of P. warrenensis with Parapliosaccomys oregonensis Shotwell from the late Hemphillian McKay Reservoir Local Fauna of Oregon. The small Perognathus is most similar to P. gidleyi Hibbard from the early Blancan, Rexroad Fauna, but the temporal significance of this comparison is uncertain. May (1981b) considered the hypsodont cricetine Repomys gustelyi to be more primitive than R. maxumi that is known from the early Blancan Maxum Local Fauna in California and from early Blancan sediments in the San Timoteo badlands (May and Repenning 1982; Albright 1999). R. gustelyi has been identified from a single tooth in the San Timoteo section associated with sediments correlated with late Hemphillian taxa (Albright 1999). Voorhies (1990) includes R. gustelyi in faunal lists from both Santee and Devil's Nest Airport Local Faunas in Nebraska, both of which are considered latest Hemphillian. Tedford et al. (2004) list Repomys (R. gustelyi) as one of the defining taxa for the late late Hemphillian (Hh4). Postcopemys valensis is known from the Hemphillian of Oregon (Shotwell 1967). The two species of Peromyscus are of little value for determining age although P. sp. A is similar to P. kansasensis from the Blancan Rexroad Fauna (Hibbard 1941). Jacobsomys has been previously identified from the early Blancan Verde Valley Local Fauna, Arizona (Czaplewski 1987). J. verdensis is also present in the early Blancan Maxum Local Fauna (Lindsay and Czaplewski 2011). Jacobsomys dailyi represents a range extension of this genus into the late Hemphillian. The three large carnivores in the Warren Local Fauna are widespread in late Hemphillian faunas of North America. The Borophagus is similar to B. secundus, which has been identified from a large number of faunas from southern and southwestern United States to Honduras. The bear, Agriotherium, is most commonly found in late Hemphillian (Hh3 and Hh4) age faunas in North America (Tedford et al. 2004). Agriotherium is thought to have been replaced by the Eurasian immigrant Ursus in faunas of Blancan age. More recently, Samuels et. al., (2009) report Agriotherium from sediments of Blancan age in Idaho, however the material is fragmentary and taxonomic identity is unclear. The saber toothed cat, Machairodus coloradensis, is known from Wikieup, Coffee Ranch, Wray, Aphelops Draw, and Edson. Machairodus probably immigrated to North America at the same time as Agriotherium (Tedford et al. 2004), but its range extends into the Blancan. The gomphothere, Rhynchotherium sp. cf. R. edensis, is very similar to R. edensis from the late Hemphillian Mt. Eden Local Fauna (Frick 1933). The shorter enamel bands on the lower tusks suggest that Rhynchotherium from Warren may be slightly more advanced than R. edensis although the two adult lower tusks are from a single individual and intra-population variability of this character is unknown. Pliomastodon vexillarius is also present in the Mt. Eden Local Fauna and is known elsewhere from faunas of late Hemphillian to late Blancan age (Madden 1980). The horse, Dinohippus edensis, is known from the late Hemphillian Mt. Eden Local Fauna (Frick 1921; MacFadden 1998). The Teleoceras material from Warren is very similar to T. cf. T. fossiger reported by Stirton (1939) from the Pinole Tuff Local Fauna of late Hemphillian age. However, Teleoceras is a common rhino in North America throughout much of the Miocene (Prothero 1998). Megatylopus ranges from the late Clarendonian to the early Blancan in the western USA (Honey et al. 1998). Both Megatylopus cf. M. matthewi and Hemiauchenia vera are known from the Coffee Ranch Local Fauna, the San Juan and Rak Camel Quarries of the Chamita Formation and the Wikieup Local Fauna all of which are late Hemphillian faunas. Hemiauchenia vera is also known from the Ogallala Group, Nebraska (including Santee and Devil's Nest Airstrip Local Faunas); Ogallala and Ash Hollow Formations, Kansas; Upper Bone Valley Fauna, Florida; Yerington, Nevada; Modesto Reservoir Local Fauna, California, and the Ocote Local Fauna, Mexico (Honey et al. 1998). The small indeterminate camelid seems to be more derived than other known Hemphillian forms. The lack of P2 and P3 parallels the morphology of the Pleistocene to recent genus Camelus. Faunal comparisons suggest a late Hemphillian age for the Warren Local Fauna with close similarities to Mt. Eden, Pinole, Yepomera, and Santee/Devil's Nest Airport Faunas. At the generic level, the Warren and Mt. Eden Faunas share Hypolagus, Repomys, Agriotherium, cf. Prosthennops, Dinohippus, Rhynchotherium, and Pliomastodon. At the specific level they share H. edensis, R. gustelyi, D. edensis, and P. vexillarius. The Warren Local Fauna and Pinole Tuff Local Fauna share Machairodus, Borophagus, Dinohippus, and ?Sphenophalos. The Warren Local Fauna shares Dinohippus, Machairodus, Agriotherium, cf. Prosthennops, and Teleoceras with the Yepomera Local Fauna of Mexico. The Warren Local Fauna shares Hypolagus, Spermophillus, Perognathus, Repomys gustelyi, Borophagus, Agriotherium, Machairodus, Dinohippus, Teleoceras, and Hemiauchenia cf. H. vera, with the Santee/Devil's Nest Airstrip Faunas of Nebraska (Voorhies 1990). Tedford et al. (2004) showed the Warren Local Fauna correlative with other late late Hemphillian (Hh4) Faunas including the Mt. Eden Local Fauna (California), the Washoe Local Fauna (Nevada), the Etchegoin sites (California), the Pinole Local Fauna (California), the Santee Local Fauna (Nebraska), the Christian Ranch Local Fauna (Texas), the Yepomera Local Fauna (Mexico), the Ocote Local Fauna (Mexico), and the Palmetto Local Fauna of Florida. The Mt. Eden Local Fauna is from reversed polarity sediments interpreted as C3r (5.23 – 5.89 Ma) (Albright 1999), the Washoe Local Fauna is overlain by a tuff dated at 4.96 +/- 0.02 (Lindsay et al. 2002), the Etchegoin sites are overlain by the Lawlor Tuff (4.83 +/- 0.04 Ma), the Pinole Local Fauna is associated with the Pinole Tuff dated at 5.2 ± 0.1 Ma (Evernden et al. 1964; Sarna-Wojcicki 1976), the Santee Local Fauna is overlain by the Santee Ash dated at 5.0 ± 0.2 Ma, the Yepomera Local Fauna is from reversed polarity sediments interpreted as Chron 3n.3r (4.896 – 4.997 Ma) (Lindsay et al. 2002). Prothomomys warrenensis n. gen. n. sp., Jacobsomys dailyi n. sp., Camelidae gen. et. sp. indet., Peromyscus sp. A, and possibly Rhynchotherium cf. R. edensis, represent morphologically advanced forms relative to known late Hemphillian taxa and are consistent with a late late Hemphillian age for the Warren Local Fauna associated with reversed polarity sediments correlated with Chron 3n.3r and overlain by the Lawlor Tuff dated at 4.83 ± 0.04 Ma. The Lawlor Tuff is the stratigraphically lowest normal polarity site in the sampled polarity sequence in the Horned Toad Formation and the 4.83 ± 0.04 Ma Ar40/Ar39 date is consistent with Chron 3n.3n (Sidufjall) (4.799 – 4.896 Ma, Lourens et al. 2004). In addition to many other known occurrences in California and Nevada, the Lawlor Tuff has also been identified at the top of the Etchegoin Formation in the Kettleman Hills, California, just above the Etchegoin Sites fauna of latest Hemphillian age (Hh4, Tedford et al. 2004). The age of the Warren Local Fauna and, by association, Member Two of the Horned Toad Formation is approximately 4.85 - 5.0 Ma. If the Hemphillian–Blancan boundary can be recognized in the Horned Toad Formation, it is likely younger than the Lawlor Tuff (4.83 ± 0.04 Ma). |
|