|
|
|
SYSTEMATIC PALEONTOLOGYThe occurrences of the taxa by locality are presented in Table 1. Synonomy of previous locality names to this work are shown in Table 2.
Class MAMMALIA Linnaeus, 1758
Cormocyon copei
Wang and Tedford, 1996
Localities. WC-1 ,WC-2.
A fragmentary P4 (JODA 8942) has no distinct parastyle. The anterolingual cingulum shows a distinct protocone.
The m1s (UWBM 39225, UWBM 40738, and JODA 8945) have a talonid basin formed by a crest-like hypoconid and enlarged entoconid. The trigonid is elongate and opened by a longitudinally oriented paraconid blade. The metaconid is prominent and projects lingually from the protoconid. The m2 has enlarged metaconids relative to the protoconid. Discussion. The inflated entotympanic bullae place the skull within the Canidae. The ventrally projected paroccipital process is not found in Hesperpocyon or the borophagines Archaeocyon, Rhizocyon, or Otarocyon, and the sagittal crest differs from the parasagittal crest condition of Cynarctoides. The paroccipital process does not fuse with the entotympanic, as in Mesocyon, Enhydrocyon, and Parenhydrocyon. Some hesperocyonines, canines, and primitive borophagines lack a parastyle on the P4 (Wang 1994; Wang et al. 1999). The size is similar to Hesperocyon gregarius, "Mesocyon" temnodon, Phlaocyon , and Cormocyon. The tooth lacks the well-developed anterior cingulum found in "Mesocyon" temnodon and H. gregarious, and the metacone blade is not shortened as in Phlaocyon. The talonid basin of the m1 and tall metaconid on the m2 are characteristic of borophagine and canine canids (Albright et al. 2008). The distinct accessory cusps on premolars and absence of diastems distinguish the dentaries from canines and the elongate trigonid differs from Archaeocyon and Rhizocyon. The lower molars lack a protostylid, which is found on Phlaocyon and Cynarctoides, and the latter genus has a more slender horizontal ramus. The crest-like talonid cusps on the m1 differ from Desmocyon and later borophagines. The sizes of the teeth are at the large end of the ranges for Cormocyon copei and larger than Cormocyon haydeni (Wang et al. 1999, their Appendix III). Geographic occurrence. Wang et al. (1999) identified C. copei from the Turtle Cove Member of the John Day Formation, and specimens questionably assigned to C. copei, have been recovered from the medial Arikareean of Colorado and late Arikareean of Florida (Wang et al. 1999; Munthe 1998).
Genus Enhydrocyon Cope, 1879
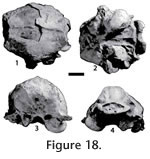
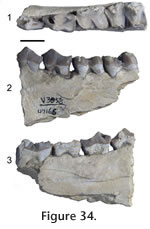
Locality. WC-2. Description. The premolars of JODA 8948 have a transversely widened base with no diastema between. The anterior two teeth are imbricated and oriented from posterolingual to anterobuccal. Matrix anterior to the p2 preserves a mold of the canine. The dentary is broken along the posterior part of the canine root, which runs nearly to the base of the dentary. The principal cusps are slightly anteriorly placed and increase progressively in height from the p2 to p4. The p2 has ridges that run along the midline from the distinct anterior cingular cusp, to the principal cusp and posterior cingular cusp. Distinct posterior accessory cusps and posterior cingular cusps form along the midline of the p3 and p4. The accessory cusps are more prominent on the latter tooth, which has a distinct anterior cingular cusp. The posterior portion of the preserved dentary is broken along the m1 alveolus. Discussion. The tooth positions are identified by the posterior position relative to the canine, indicated by an alveolus and posterior enamel a tooth larger than the p2, and the anterior position relative to the m1, indicated by the alveolus postero-lingual of the p4. Absence of the p1, large premolars that are wide, and no diastemas are diagnostic characters of Enhydrocyon and Philotrox. Philotrox retains the p1 and is much smaller (Wang 1994). Enhydrocyon basilatus is much larger than this specimen, and E. crassidens (Wang 1994). Specimen JODA 8948 shows the presence of accessory cusps as in Enhydrocyon pahinsintewakpa and Enhydrocyon stenocephalus; however, premolar morphology and size do not distinguish these species (Wang 1994). Molar and skull morphology characters not preserved in specimen JODA 8948, distinguish E. pahinsintewakpa from E. stenocephalus. Geologic occurrence. Enhydrocyon is known from the Arikareean of the Great Plains and Oregon. Enhydrocyon pahinsintewakpa and E. stenocephalus are confined to the early Arikareean of the Great Plains and Oregon, respectively.
Genus Parenhydrocyon Wang, 1994 Referred material. UWBM 39226, partial right dentary with p4 and m1: dental measurements in Table 3. Locality. WC-2. Description. The alveolus of the p3, and the p4 and m1 are present with no diastemata in UWBM 39226. The p4 has a worn principal cusp, and the m1 has a worn paraconid blade and protoconid. The p4 is slender with a distinct posterior accessory cusp and posterior cingular cusp. The trigonid of the m1 is dominated by the protoconid, but has an unreduced metaconid. The talonid is trenchant and dominated by the hypoconid. The entoconid forms a wide shelf. Discussion. The trenchant talonid on the m1 differs from the basined condition in the amphicyonid Daphoenus, and is present in the amphicyonids Daphoenictis and Brachyrhynchodon and in hesperocyonine canids (Hunt 1998; Wang 1994). The narrow p4 is diagnostic of the hesperocyonine Parenhydrocyon (Wang 1994; Munthe 1998). The metaconid in Parenhydrocyon jospehi is not reduced as in Parenhydrocyon wollovianus and Parenhydrocyon robustus (Wang 1994). The p4 retains accessory cusps, which are variable in P. jospehi (Wang 1994). Geologic occurrence. Parenhydrocyon josephi is known from the Whitneyan through the Arikareean of the Great Plains and Oregon.
Order RODENTIA Bowditch, 1821
Locality. WC-3. Description. A fragment of a lower incisor, specimen UCMP 119462, contains enamel restricted to the anterior surface. Approximately one-half of the enamel surface is smooth, and three lateral ridges run longitudinally along the tooth. The tooth measures 1.9 mm wide. Discussion. Longitudinal ridges on incisors are present in several families of rodents in the North America Paleogene (Korth 1998), but three small and one large ridge are found on the lower incisors of Paciculus and Leidymys (Lindsay 2008). Geologic occurrence. Both genera first appear in Oregon in the late early Arikareean (Albright et al. 2008). In the Great Plains, these taxa occur earlier than Oregon where Leidymys and Paciculus are known as early as 30 Ma and extend into the Miocene.
Order LAGOMORPHA Brandt, 1855
Locality. WC-3. Description. The bone on the specimen is badly weathered and broken. The incisors are shattered. A partial dentary preserves three molariform cheek teeth and a p3. The teeth are hypsodont. The enamel is thicker on the posterior and buccal sides of the trigonid and talonid. A thin band of enamel connects the trigonid and talonid lingually. The right p3 was prepared down to the base to show an occlusal cross section, where the bone and enamel are better preserved. An external reentrant extends one-third across the tooth and separates the trigonid from the talonid. The long axis of the reentrant is transverse and not oriented obliquely. Discussion. Whereas the dentary is poorly preserved, the p3 has at least one external reentrant. The overall shape of the p3 is similar to Archaeolagus as figured by Dice (1929, Figure 1) and described by Dawson (1958, 2008), but differs in lacking an anteroexternal reentrant that persists during wear. The internal reentrants of p3s in palaeolagines disappear during heavy wear (Dice 1929). Only the base of the tooth is preserved, representing a heavily worn p3. The absence of the anteroexternal and internal reentrants is consistent with a well worn p3 of a palaeolagine leporid. The cheek teeth are more hyspodont than Mytonolagus and smaller than Megalagus described by Dawson (1958 table 2). The p3 is not reduced as in Desmatolagus and not shortened as in Chadrolagus (Dawson 1958, 2008). Palaeolagus intermedius, Palaeolagus philoi, and Palaeolagus? sp. are similar in size and lack persistent internal reentrants. Characters distinguishing P. intermedius and P. philoi are in upper cheek-tooth morphology (Dawson 1958). Geologic occurrence. Palaeolagus is known from the Chadronian to the early Arikareean of the Great Plains, the Chadronian of Saskatchewan, Whitneyan of California, and early Arikareean of Texas and Oregon (Prothero and Emry 2004; Tedford et al. 2004).
Order PERISSODACTYLA Owen, 1848
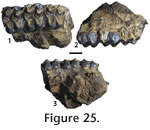
Locality. WC-2. Description. Distinct paraconules and metaconules form on the protoloph and metaloph of the P2 and the left P or M. The P2 is identified by an anteriorly positioned parastyle. The two other teeth are not distinguishable between molars or premolars because of the molariform premolars in equids. The protoloph connects to the ectoloph. A spur on the lingual portion of the ectoloph projects to, but does not connect to, the metaloph. A strong intervallic cusp is on the P2. A weak intervallic cusp is seen on the P or M. The anterior cingulum is strong on the P or M. A small pocket forms between the anterior portion of the hypostyle and the posterior cingulum. The ectoloph is partially preserved on the P or M and shows a shallower W-shaped morphology. Discussion. The distinction between Miohippus and Mesohippus is not clear (Prothero and Shubin 1989; MacFadden 1998). MacFadden (1998) stated that a comprehensive reevaluation of the species of Mesohippus and Miohippus is needed. Prothero and Shubin (1989) reevaluated several species of these two genera from the Great Plains, but drew no conclusions about many species outside of that geographical area. Prothero and Shubin (1989) used the size of the hypostyle to distinguish between Mesohippus and Miohippus. The referred teeth display large class 3 hypostyles, larger than those of any species of Mesohippus. The size of the teeth is slightly larger than ?Miohippus equinanus, the smallest species of Miohippus, and smaller than Miohippus obliquidens measured by Prothero and Shubin (1989, table 10.2), but metaconules are more pronounced, and the metaloph and protoloph are less obliquely oriented than in the latter species. The prominent paraconules and metaconules distinguish JODA 8946 from Miohippus acutidens, Miohippus anceps, and Miohippus equiceps described and figured by Osborn (1918). The large class 3 hypostyles, prominent parastyle on the P2, small size relative to other species of Miohippus shown in specimen JODA 8946 best fit ?M. equinanus (Osborn 1918; Prothero and Shubin 1989). JODA 8948 is much smaller than Kalobatippus (Osborn 1918). Osborn (1918) placed ?M. equinanus questionably in Miohippus as it features an advanced dentition for a Mesohippus sized equid, but is only known from dental material. Geologic occurrence. ?M. equinanus is known from the Whitneyan to early Arikareean of South Dakota, and the Whitneyan of Oregon (MacFadden 1998).
cf. Miohippus equiceps
(Cope), 1878
Locality. WC-2. Description. Three heavily worn teeth have enlarged paraconules and metaconules that are distinct on the metaloph and paraloph, respectively. The anterior portions of the parastyles are missing. The metaloph connects to the ectoloph on P3. Lingual cingulum is weak and forms a weak intervallic cusp. The anterior cingulum is strong and forms a tubercle. The metaloph connects to the ectoloph. The hypostyles are triangular and represent at least class 2 hypostyles (Prothero and Shubin 1989). The anterior portion of the enamel on the ectoloph is preserved and suggests a W-shaped morphology. Discussion. The size of teeth, distinct paraconules and metaconules, intervallic cusps, metaloph connecting to the ectoloph, and large triangular hypostyles are similar to Miohippus equiceps (Osborn 1918; Prothero and Shubin 1989). Miohippus annectans and M. gidleyi are larger species (Prothero and Shubin 1989; Osborn 1918), and the teeth differ from similar sized M. obliquidens and M. assiniboiensis in having prominent metaconules and strong lingual cingula. Geologic occurrence. Miohippus equiceps is known from the Whitneyan and early Arikareean of Oregon, and early Arikareean of the Great Plains (Osborn 1918; MacFadden 1998).
Miohippus sp.
Localities. WC-2, WC-3 Description. The lower cheek teeth are molariform. The cingulum is continuous along the hypoconid and the protoconid. The paralophid is continuous with the metastylid. The metaconid and metastylid are indistinct and form a high pillar. The paralophid and hypolophid do not connect.
Family RHINOCEROTIDAE Gray, 1821
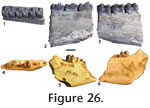
Localities. WC-1, WC-2. Description. The molariform teeth have a simple square-shaped loph morphology. A weak crochet is present. The buccal enamel is rugose, and the anterior and lingual cingula are strong. A complete tooth (JODA 8949) is 27.1 mm long and 38.9 mm wide. A partial tooth is missing the buccal portion of the ectoloph, and the metaloph and protoloph are worn to the point that they unite. Discussion. The simple square-shaped loph, strong cingula, and size place these specimens within Diceratherium annectens (Prothero 2005, table 4.4). Geologic occurrence. D. annectens is known from the Arikareean of the Great Plains, early to middle Arikareean of Oregon, and the late Arikareean of Texas (Prothero 1998b, 2005). Diceratherium sp. Referred material. JODA TS070510, partial left P or M. Locality. WC-3. Description. An upper left cheek tooth preserves a nearly complete metaloph and anterior hypoloph. The enamel is rugose. The anterior cingulum is strong and runs buccally. A cuspid projects posteriorly from the posterior portion of the metalophid. Discussion. The large tooth exceeds the size of D. annectans and approaches D. armatum (Prothero 2005), but accurate measurements are not possible.
cf. Diceratherium sp.
Locality. WC-2. Description. A lower premolar (JODA DG06051) includes a complete metalophid and the anterior portion of the hypolophid. The metalophid and hypolophid do not connect. The anterior cingulum is weak. The specimen measures approximately 21.3 mm wide. The lower molariform tooth (JODA TS070516) has a nearly complete protolophid and partial metalophid. The protolophid shows a simple L-shaped morphology. The tooth is high crowned, and the enamel is rugose. Discussion. The loph pattern and size indicate Diceratherium (Prothero 2005).
Family HYRACODONTIDAE or RHINOCEROTIDAE
Locality. MC-1. Description. The specimen preserves a nearly complete L-shaped metalophid. The metalophid is 21.3 mm wide. Posterior cingulum is present. Discussion. This specimen was collected as float by D.A. Swanson from the Milk Creek tuff, south of Kloochman Rock, and is the lowest stratigraphic occurrence of a terrestrial vertebrate in the Tieton River area (Swanson 1964, 1978). L-shaped metalophids are characteristic of hyracodontids and rhinocerotids, both present in the Eocene and Oligocene of North America (Prothero 1998a, 1998b).
Order ARTIODACTYLA Owen, 1848
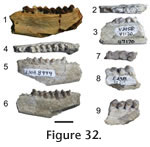
Locality. WC-2. Description. The partial dentary has a complete m2 and roots of p4, m1, and m3. The m2 is slightly worn, rectangular in outline, and bunodont. Four primary cusps are present and nearly identical in size. The protoconid and metaconid tips are slightly higher than the tips of the hypoconid and entoconid. A small hypoconulid occurs on the posterior cingulum. A cuspid lies in the anterior portion of the valley between the hypoconid and entoconid. The anterior cingulum is weak. The buccal cingulum is present but does not continue along the metaconid and hypoconid. Discussion. The m2 is identified on the basis of size relative to adjacent roots. The rectangular outline of the molar, the bunodont condition of the tooth, with four nearly subequal principal cusps is characteristic of tayassuids (Pearson 1923, Wright 1998). The m2 is identical in cusp morphology to the lower m2 of Cynorca cf. C. sociale illustrated in Woodburne (1969, Figure 3) and of Perchoerus described and figured by Pearson (1923, Figure 17). Woodburne (1969) described the species C. sociale as similar to Perchoerus, but stated that the lower dentition is conservative in tayassuids. This specimen could represent either, but Perchoerus is known from the Whitneyan and early Arikarrean of the Great Plains and Oregon.
Family HYPERTRAGULIDAE Cope, 1879
Localities. WC-2, WC-3. Description. The dentaries have closely spaced cheek teeth. The p4 in specimen UWBM 29151 shows a prominent paraconid and metaconid, which form a crest extending posterior to a subcircular hypoconid. The lower molars are selenodont and have crescent-shaped cuspids. A cingulum is present on the anterior portion of the molars, located between the anterior and posterior selenes, and forms a small intervallic cuspid. Discussion. These specimens are assigned to Hypertragulidae by the small size, selenodont dentition, and prominent paraconids and metaconids on the p4s. The strong cingula and cuspids, and low crowns allow assignment to Hypertragulus and distinguish these specimens from Nanotragulus (Webb 1998; Metais and Vislobokza 2007). Geologic occurrence. Hypertragulus is known from the Whitneyan to the late early Arikareean of Oregon and the Great Plains (Webb 1998).
Family MERYCOIDODONTIDAE Hay, 1902
Localities. WC-2, WC-4. Description. The teeth are selenodont. A partial left dp4 preserves the median and posterior lobes and the partial right dp4 preserves the posterior lobe. The selenes are rotated so that the apex of the crests and crescents point slightly posterior. The anterior crescents of the cheek teeth have strong mesostylids that overlap but do not connect to the anterior part to the posterior crescents. The metastylids overlap the anterior portions of the following tooth's anterior crescent. Parastylids are absent. The anterior and posterior crests do not meet, and are divided by a deep valley. A cingulum is present on the anterior portions of the teeth and between the anterior and posterior crests. The dorsal surface of the mandible widens at the m2. The anterior, median, and posterior crests are strong on the p3 and p4. The posterior intermediate crest is prominent and extends from the middle of the median crest and runs parallel to the posterior crest. The anterior and posterior crescents are weak. The posterior basin on p3 is not closed, and a prominent posterior intermediate crest is parallel to the posterior crest. Discussion. Recent overviews of the higher level taxonomy of oreodonts are discussed in Lander (1998) and Stevens and Stevens (1994, 2007). The identifications of these specimens follow earlier generic and specific descriptions by Thorpe (1937) and Schultz and Falkenbach (1949, 1968) and these are placed within the larger taxonomic framework of Stevens and Stevens (2007). All specimens identified as oreodonts, described above and below, show selenodonty that exclude these as agriochoerids. None of the oreodont specimens from the Wildcat Creek beds shows the hypsodont condition or the shallow valleys separating selenes in the molars of leptauchineine oreodonts. Enamel is thicker in these specimens than in Miniochoerus. Two specimens preserving lower dentition from the recent collections are referred to Eporeodon. These specimens were compared with Eporeodon in the UCMP collections and examples figured and described by Loomis (1924), Thorpe (1937), and Schultz and Falkenbach (1968). In particular, the p3 characterizes Eporeodon in having prominent posterior intermediate crests on the premolars that extend posteriorly from the median crest and divide an open posterior basin. These specimens are smaller than Mesoreodon. The molars are not as transversely widened as in Merycoidodon (Stevens and Stevens 1994). The premolars have stronger crescents and posterior intermediate crests than Merycoides, Paroreodon, and Phenacocoelus. The premolars of these specimens do not show the degree of molarization as Ticholeptus. The premolars are not anteroposteriorly shortened as Paroreodon, Oreodontoides, Merychyus, and Paramerychyus (Schultz and Falkenbach 1947; Stevens and Stevens 2007). These specimens represent oreodonts smaller than Desmatochoerus, Promerycochoerus, Megoreodon, Hypsiops, Merycochoerus, and Submerycochoerus. Geologic occurrence. Eporeodon is known from the Whitneyan through the early Arikareean of the Great Plains, California, and Oregon (Stevens and Stevens 2007).
Genus Merycoides
Douglass, 1907
Locality. WC-2. Description. The occlusal enamel of the p3 and m2 is nearly worn through. Anterior, median, and posterior crests are present on the p3, but the median crest is mostly broken. The posterior intermediate crest extends from the median crest and runs parallel to the posterior crest. The posterior crescent is reduced. The p4 has more prominent anterior, median, and posterior crests, lacks an anterior crescent . The posterior intermediate crest projects from the median crest, is reduced, and does not connect to the posterior cingulum. The apex of the crests and crescents of the m1 and m2 project posteriorly. The anterior and posterior crests are divided by a deep valley; the cingulum is strong in the valley. Discussion. This specimen is referred to Merycoides because it has anterioposteriorly shortened premolars, reduced crests on the p3, and posterior and posterior intermediate crests on the p4 that form an undivided posterior basin (Thorpe 1937, Stevens and Stevens 2007). Anteroposteriorly shortened premolars are found in merychyines (Stevens and Stevens 2007), but UCMP 47166 lacks the tooth row offset, as in Oreodontoides and Phenacocoelus (Schultz and Falkenbach 1947). UCMP 47166 retains the labial cingula, unlike Merychyus (Stevens and Stevens 2007). The tooth row is more crowded than Paramerychyus and Paroreodon (Schultz and Falkenbach 1947). This specimen differs from ticholeptines in lack of elongate premolars (Stevens and Stevens 2007). Geologic occurrence. Merycoides first occurs in the John Day Formation and is known from North America from the latest Whitneyan through the Arikareean (Tedford et al. 2004; Stevens and Stevens 2007).
Genus Mesoreodon Scott, 1893
Localities. WC-2, WC-4. Description. The p3, dp4, and m1 (JODA NT06052N) show light wear. A partial p3 preserves a strong anterior crest. The anterior, median, and posterior lobes of the dp4 are rotated so that the apices of the crests and crescents point posteriorly. The posterior portion of each crescent overlaps the anterior portion of the following crescent. The anterior and posterior crests do not join and are divided by a deep valley. The p3 has prominent anterior, median, and posterior crests. The median crest directs posterior and lingually from the primary cusp, and joins with the posterior crescent. The posterior intermediate crest is prominent and extends from the posterior cingulum and partially divides the posterior basin.
Discussion. Specimen JODA NT06052N was collected in situ in Thunder Creek and represents the highest such occurrence, 228 m above the pumice lapilli-tuff. Specimens are referred to Mesoreodon by having reduced anterior intermediate crests on the upper premolar, and prominent posterior intermediate crests and crescents on the lower premolars (Loomis 1924; Thorpe 1937; Shultz and Falkenbach 1949). These specimens are larger, have more prominent crescents on the lower p3, and have posterior intermediate crests that originate from the posterior cingulum, not the median crest as in Eporeodon. The prominent posterior intermediate crest on the premolars differs from Promerycochoerus and Hypsiops (Shultz and Falkenbach 1949). These teeth represent individuals smaller than Desmatochoerus and Merycochoerus (Stevens and Stevens 2007). The premolars are not reduced as in Merycoidodon, Merychyus, Submerycochoerus, and Paroreodon, and are not rotated as in Oreodontoides, Phenacocoelus, Paramerychyus, and Ticholeptus (Schultz and Falkenbach 1941, 1947; Stevens and Stevens 2007). The tooth row is not crowded as in Merycoides and Megoreodon (Schultz and Falkenbach 1954). Geologic occurrence. Mesoreodon first appeared in the Chadronian of Texas, and is known from the Chadronian through the early Arikareean of the Great Plains and the early Arikareean of Oregon (Stevens and Stevens 2007).
Genus Promerycochoerus
Douglass, 1901
Localities. WC-3, WC-4. Description. Specimen UCMP 128070 includes the right M2 and M3 tooth roots, partial right jugal, postglenoid process, basioccipital region, and foramen magnum. The snout, most of the palate, occipital condyles, auditory bullae, and the dorsal skull are missing. The postglenoid processes are heavy. The jugal is robust and relatively flat on the lateral side. The preserved tooth roots are large. The M3 metastyle is the only preserved portion of the dentition, and it is prominent and posteriorly projected.
Specimen JODA TS070523 includes the frontal region to the postorbital constriction and has a partial right and left M2 and M3. The skull is dorsoventrally flattened, and the right buccal crests are deformed lingually. The preorbital fossa is broad and shallow. The jugals are broad, and the frontals are wide. The teeth are selenodont. The apex of the crests and crescents projects anteriorly. The anterior and posterior crescents are divided by a valley, and do not meet. The lingual cingulum is strong and forms a prominent intervallic cusp on the M3. The styles and ribs are prominent on the M3 and the metasyle projects buccally from the tooth. A large partial P4 is indicated by the undivided selene.
These specimens, taken together, represent Promerycochoerus superbus, based on the prominent and robust postglenoid processes, paroccipital processes prominent, fan-shaped occiput, extension of the palate posterior to the M3, and strong external ribs on the molars (Thorpe 1937; Schultz and Falkenbach 1949). The specimens are substantially larger than Mesoreodon and Desmatochoerus and differ from Ustatochoerus by having a pronounced facial vacuity and lower crowned teeth (Stevens and Stevens 2007). The large, wide, robust, postglenoid processes differ from Megoreodon and P. [Paramerycochoerus] barbouri, but are not as robust as Promerycochoerus carrikeri and Promerycochoerus chelydra described and figured by Schultz and Falkenbach (1949, 1954). Geologic occurrence. Promerycochoerus superbus occurs from the late early and early late Arikareean of South Dakota but first occurs at the Deep Creek Tuff, dated at 27.8 Ma, of Oregon (Janis et al. 1998; Stevens and Stevens 2007).
Merycoidodontidae genus and species
Locality. WC-1 Description. A partial pelvis includes complete right acetabulum and parts of the ilium, ischium and pubis. The base of the ilium is wide, but no ridge is preserved. A partial maxilla preserves the anterior portion of the zygomatic arch. The teeth are from a large, Promerycochoerus superbus-sized oreodont. The zygomatic arch has rodent gnawmarks 8 mm long and about 0.5 mm wide. Discussion. Specimen JODA TF10031 is the lowest in situ occurrence of a fossil in the Wildcat Creek beds, 120 m above the pumice lapilli tuff. Rodent gnawing on specimen UCMP 47165 indicates the fossil was exposed prior to burial. |
|