|
|
|
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linnaeus, 1758 Type Species. Megapeomys lavocati Fejfar, Rummel, and Tomida, 1998, by original designation.
Megapeomys repenningi sp. nov.
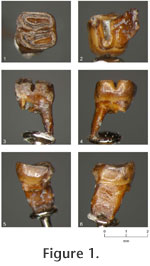
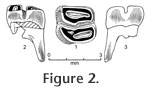
1998 - Megapeomys sp. Fejfar, Rummel, and Tomida, p. 138, figure 9. Holotype: Isolated left lower first molar (m1) (NMNS-PV 20153). NMNS stands for the National Museum of Nature and Science in Tokyo, Japan, which was formerly called the National Science Museum. Etymology: To the honor of the late Charles A. Repenning, for his great contribution to the paleontology of rodents and other mammalian groups. Type locality and age: Exposures on the left bank of Kiso River near Dota, Kani City, Gifu Prefecture, central Japan (Tomida and Setoguchi 1994, figure 1-1), near the top of Nakamura Formation; likely around 19 Ma (21 – 18 Ma is greatest probable range) and is correlated with MN 3 zone of European land mammal age (see Geology and Age section above). Diagnosis: Relatively small-sized species of the genus. Lower first molar with synclinid III deeper; lingual end of the synclinid III becomes shallower only slightly; distal root of m1 does not split toward the tip.
Description: Relatively deeply worn m1, barrel-shaped and relatively high crowned (for an eomyid). Mesial roots are broken at the base and not securely known for number and shape. Distal root is single with mesio-distally compressed plate-like shape, becoming only slightly narrower toward the tip, and not splitting toward the tip. The enamel is rather thick as seen on the occlusal surface. The occlusal surface is concave, and enamel wall on the lingual side is higher than buccal side. The crown consists of two, laterally elongated oval lobes separated by a deep transverse valley (synclinid III), which becomes shallower only slightly toward the lingual end. Synclinid III is almost transverse, but in detail it bends rather sharply at a point somewhat buccal to the midpoint of the width. Mesial lobe is slightly narrower than distal lobe, but is about the same in mesio-distal length. Its central valley (synclinid II) is as wide as it can be relative to the width of mesial lobe and is nearly transverse, only slightly diagonal to the mesio-distal direction of the tooth. Distal lobe is slightly wider than mesial. Its central valley (synclinid IV) is short, only about two thirds of its maximum possible width, and is skewed lingually. Beyond where the synclinid ends, a small enamel island is present on the buccal one third of the distal lobe.
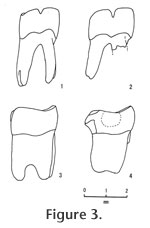
M. repenningi is slightly larger (10%) than M. lavocati and is smaller than M. lindsayi, also from Bohemia. Although M. lindsayi is represented only by p4 (length 2.53 mm, width 2.50 mm in holotype), it is about 20% larger than M. lavocati (length 2.00 mm, width 2.17 mm in holotype; Fejfar et al. 1998). The height of the lingual edge of synclinid III above the enamel base is about 40% of tooth length in M. repenningi, while it is nearly one half in M. lavocati, suggesting that synclinid III is deeper at its lingual end in M. repenningi than M. lavocati (Figure 3). Distal roots of m1 and m2 in M. lavocati are split into two at approximately the lower one third of the root, unlike in M. repenningi (Figure 3; figure 7 of Fejfar et al. 1998). This may suggest that M. repenningi is somewhat more primitive than M. lavocati, because the same root is always single in Apeomys, the ancestral form of the genus Megapeomys (Fejfar et al. 1998). Although the genus Megapeomys was originally known from Europe (Czech Republic) and Asia (Japan), another species of the genus (M. bobwilsoni; Morea and Korth 2002) and a similar but derived genus (Apeomyoides savagei; Smith et al. 2006) were recently described from North America. In A. savagei, all cheek teeth are higher crowned than Megapeomys in general and moreso than M. repenningi in particular, although the size is similar to M. repenningi. M. bobwilsoni is the largest species of the genus and has four roots on m1-2, while M. repenningi is a much smaller species and has three roots on m1. Thus, M. repenningi can be easily distinguished from apeomyine species from North America. |
|