|
|
|
SYSTEMATIC PALEONTOLOGY
Class MAMMALIA Linneaus, 1758 Comments. Recent analyses of the suprafamilial relationships of the "Triisodontidae" within the Placentalia have been inconclusive. The family is left incertae sedis within the cohort. Following Van Valen (1978) Eoconodon coryphaeus is recognized as the type species of the genus. Biogeographically, the species of Eoconodon fall into two groups, those known only from the San Juan Basin and possibly Texas, and those known only from localities in Wyoming, Montana, and Saskatchewan. To facilitate this study the type and other species from the San Juan Basin (E. gaudrianus and E. ginibitohia) as well as possible occurrences in Texas are briefly reviewed. Then records of Eoconodon from localities in the northern Western Interior, including descriptions and analyses of new material from Montana, are presented. Species known from the southern Western Interior, New Mexico, and possibly Texas Eoconodon coryphaeus (Cope, 1885) Comments. Eoconodon coryphaeus, the largest known species of genus, is represented by large samples from the Hemithlaeus kowalevskianus - Taeniolabis taoensis Zone (Pu2) and smaller samples from the Taeniolabis taoensis - Periptychus carinidens Zone (Pu3) of the San Juan Basin (Williamson 1996). The species has yet to be certainly identified from localities outside the San Juan Basin. The nomenclatorial history of Eoconodon coryphaeus is long and complex (see Matthew 1937, Van Valen 1978, Williamson 1996), but an adequate revised diagnosis of the species has yet to be developed. Remedying this situation will require a thorough analysis of the large samples now available from the San Juan Basin, a task beyond the scope of this paper. Currently the primary if not sole character usually cited to distinguish E. coryphaeus from the other species of Eoconodon is the larger size of its postcanine dentition (Table 1). For the purposes of this study, the difference in dental dimensions is deemed sufficient as a provisional diagnosis. Eoconodon gaudrianus (Cope, 1888) Diagnosis. Williamson (1996, p. 41) essentially followed Matthew (1937) and summarized the diagnosis of the species as follows: "This species differs from Eoconodon coryphaeus in its smaller size. It differs from Goniacodon in the greater transverse diameters of the molars and the unreduced M3." Comments. The mean values of the lengths of m2 and m3 (Table 1) suggest that m2 of Eoconodon gaudrianus was slightly longer than m3, but the samples are small and probably do not adequately reflect individual variation in size of the dentition. AMNH 3200, the type specimen, includes fragments of a skull, dentaries, and a calcaneum. Reasonably well preserved P4 and M3 are present in the fragment of maxillary; one dentary includes remains of m2 and m3. Stressing that little beyond the margins of the alveoli of these lower molars are preserved, the crown length of m2 (alveolar length ca. 9.0 mm) might have been greater than that of m3 (alveolar length ca. 8.8 mm), however, an allowance for a posteriorly projecting hypoconulid region of m3 must be made. Matthew (1937) and later workers appear to have placed greater weight on the morphology of the somewhat better preserved lower molars of AMNH 4029 and AMNH 58116. On the basis of this material it appears that Eoconodon gaudrianus is characterized by presence of an m3 with a length equal to or longer than that of m2. A survey of the dentitions of Protungulatum as well as Baioconodon and Oxyclaenus, which have been considered to be closely related archaic ungulate sister groups (e.g., Archibald 1998), suggests the working hypothesis that an m3 of approximately equal or greater length than m2 was the primitive condition for Eoconodon. Occurrences. In contrast to Eoconodon coryphaeus, E. gaudrianus is a rare species in Hemithlaeus kowalevskianus - Taeniolabis taoensis Zone (Pu2) local faunas of the San Juan Basin and unknown in the smaller samples of Taeniolabis taoensis - Periptychus carinidens Zone (Pu3) local faunas (Williamson 1996). The localities at which the type and other specimens from the E.D. Cope collection referred to E. gaudrianus by Matthew (1937) are not precisely recorded. Williamson (1996) noted occurrences of E. gaudrianus in three local faunas in the Hemithlaeus kowalevskianus – Taeniolabis taoensis Zone (Pu2). Eoconodon ginibitohia Clemens and Williamson, 2005 Revised Diagnosis. Size: dimensions of p4 and m2-3 either just fall within or are slightly smaller than the observed ranges of variation of homologous teeth, where known, of Eoconodon nidhoggi and are distinctly smaller than the observed ranges of variation of dimensions of E. gaudrianus, E. coryphaeus, the new species of Eoconodon described below, and probably E. copanus. The m2-3 are distinguishable from those of E. nidhoggi by: 1) greater reduction of the paraconid; 2) talonid cusps of m2 and, probably, m3 are relatively smaller and less distinct; 3) the basal cingulid is not as well developed and incomplete across the labial side of the trigonid, and 4) the talonid of m3 is narrower. In the single available specimen, length of m3 is very slightly less than that of m2, and the trigonid and particularly the talonid of m3 are narrower than those of m2 (Table 1). Comments. The presence of a new species of Eoconodon in the Hemithlaeus kowalevskianus – Taeniolabis taoensis Zone (Pu2) of the San Juan Basin, was first recognized by Williamson (1996) on the basis of a single, fragmentary mandible. (NMMNH P21622) preserving p4 and m 2-3 that was described and named E. ginibitohia by Clemens and Williamson (2005). The taxon referred to as E. cf. E. gaudrianus in the original diagnosis of E. ginibitohia is the new species of Eoconodon described below. ?Eoconodon sp. Standhardt (1986) described several fragmentary teeth and postcranial bones from LSUMG locality VL-107 ("Glen Eleven"), upper Javelina Member, Tornillo Formation and referred these to Eoconodon coryphaeus. Williamson (1996, p. 66) argued that these specimens, possibly from the same individual, are too fragmentary to allow a generic determination but suggest the presence of a large "triisodontid." Standhardt (1986, 1995) also noted the possible presence of Eoconodon in LSUMG locality VL-108, which she thought was of late Puercan age primarily on the basis of paleomagnetic evidence. On the basis of faunal composition Williamson (1996, p. 66-67) suggested a Torrejonian correlation. Additional material is needed to validate this purported occurrence. Species known from the northern Western Interior, Wyoming, Montana, and SaskatchewanEoconodon nidhoggi Van Valen, 1978 Holotype. UMVP 1471, left m3 (Van Valen 1978, plate 4, fig. 6). Type Locality. Purgatory Hill (UCMP loc. V-71202), Tullock Formation, McCone County, Montana, Pu2/Pu3 (Van Valen and Sloan 1965). Referred Postcanines and Localities. In establishing this species Van Valen (1978) did not refer any other specimens from Purgatory Hill or other localities in McCone or Garfield Counties. Casts of several isolated teeth from Purgatory Hill in the UMVP collection identified as Eoconodon sp. have been distributed to several museums. One of these, a cast of an isolated P4 (UMVP 1475), might represent a tooth referable to E. nidhoggi. The majority of the specimens currently included in the hypodigm of the species are from localities in Garfield County, Montana. M1 or M2, UCMP 156122 (V-72201), UCMP 170847, 189400 (V-73080), UCMP 156110 (V-75230); m1, LACM 32925 (V-72125), UCMP 145325 (V-73080), UCMP 192015 (V-73082); m2, UCMP 192014 (V-72129), UCMP 145328, 170848, 185983, 186602 (V-73080); m3, UCMP 145323 (V-72126), UCMP 145330, 186414 (V-73080). Revised diagnosis. As far as they are known the lengths and widths of the molars are smaller than of those of the other species of Eoconodon except E. ginibitohia. Dimensions of m2 and m3 of the type and only specimen of E. ginibitohia are either slightly smaller than or fall within the lower end of the observed ranges of variation in the small samples of homologous teeth of E. nidhoggi (Table 1). The m2 and m3 of E. nidhoggi are distinguishable from those of E. ginibitohia by: 1) relatively larger paraconids; 2) talonid cusps of m2 and, probably, m3 are relatively larger and more distinct; 3) the basal cingulid is better developed and complete across the labial side of the trigonid, and 4) the talonid of m3 is broader than its trigonid. E. nidhoggi resembles E. coryphaeus in the greater length of m3 relative to m2. In E. ginibitohia the length of m3 is slightly smaller (99%) than that of m2. In contrast, the mean length of m3 of the new species described below is distinctly shorter (77%) than that of m2. Occurrences. In addition to Purgatory Hill, the type locality (V-71202) in McCone County, specimens of Eoconodon nidhoggi have been found the following localities of Pu2/Pu3 age in the Tullock Formation, Garfield County: V-72125, V-72126, V-72129, V-72201, V-73080, V-73082, and V-75230. Specimens tentatively referred to E. nidhoggi have been reported from the following localities outside Garfield and McCone counties: Buckley (1994) identified isolated upper and lower molars from Simpson Quarry, Bear Formation, south central Montana, as Eoconodon sp. cf. E. nidhoggi. The age of this local fauna cannot be refined beyond Pu2/Pu3 undifferentiated (Buckley 1994; Lofgren et al. 2004). Noting its diminutive size Hunter et al. (1997) described a small fragmentary lower molar from the Hiatt South Locality, Fort Union Formation, southeastern Montana and tentatively referred it to E. nidhoggi. On biostratigraphic grounds Hunter et al. (1997) argued for a Pu2, possibly early Pu2, age assignment for this locality (Lofgren et al. 2004). Finally Johnston and Fox (1984) identified three fragmentary teeth found at Rav W-1, Saskatchewan, as ?Eoconodon nidhoggi. Considering the stage of evolution of some of its members Fox (1997) tentatively assigned a Pu2 age to this local fauna, but Lofgren et al. (2004) tentatively assigned it to the Pu3 interval zone on the basis of the presence of a species of Taeniolabis. Description and comparisons. Tentative identifications of the P3 and P4 of the two species of Eoconodon currently recognized in the Pu2/Pu3 of northeastern Montana are based on comparisons with the dentition of E. coryphaeus from the San Juan Basin. In the latter species the crowns of P3 and larger P4 are dominated by a large, bulbous paracone, which can be slightly canted posteriorly. On both premolars, a low protocone, relatively larger on P4, is present (see Matthew 1901, figure 12). The width of the crown of P4 is approximately the same as that of M1; P3 is not as broad. Assuming that the posterior upper premolars of E. nidhoggi are morphologically similar to those of E. coryphaeus, an isolated P4 (UMVP 1475) from Purgatory Hill (V-71202) can be tentatively referred to E. nidhoggi. Its paracone, the highest cusp, dominates the crown and is slightly canted posteriorly. Labially a weakly developed basal cingulum links the salient parastylar region and the shorter and slightly higher metastyle. These stylar cusps are approximately the same height as the diminutive protocone. Posteriorly, a weakly developed basal cingulum extends from the metastyle to the protocone. Neither conules nor a cusp on the posterior slope of the paracone are present. The width of the crown of UMVP 1475 falls within the observed range of variation of width of isolated upper molars referred to E. nidhoggi.
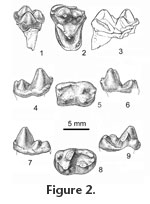
Of the species of Eoconodon, only E. coryphaeus is known from skull fragments containing both M1 and M2 with crowns sufficiently well preserved to provide data on their length/width ratios. The morphological differences distinguishing these teeth are limited. In any specimen of E. coryphaeus preserving both molars, M1 is longer anteroposteriorly, and its length/width ratio is greater than that of the associated M2. Expanding the sample to include maxillary fragments preserving either M1 or M2, it becomes apparent that the ranges of variation of the length/width ratios of these molars overlap. Analysis of this expanded sample of E. coryphaeus including 12 M1s and an equal number of M2s shows the range of the length/width ratio of the M1s is 0.71-0.93 (mean = 0.83); the range of the length/width ratio of the M2s is 0.65-0.87 (mean = 0 .75). On M1s of E. coryphaeus, the ectocingulum is divided by a shallow ectoflexus into relatively symmetrical lobes; on M2s the posterior lobe can be slightly smaller and deflected anteriorly. The differences are not great. Finally, the precingulum and postcingulum tend to be slightly broader and extend farther lingually on M1 than on M2. Because of breakage the length/width ratio for only one of the four molars, UCMP 156110, was determinable and it (0.79) lies within the overlap of the ranges of variation of M1 and M2 of E. coryphaeus. As far as can be determined the ectocingulum was divided into relatively symmetrical lobes resembling M1s of E. coryphaeus. Differences in length and breadth of the precingulum and postcingulum are greater on molars of E. coryphaeus than on the isolated teeth referred to E. nidhoggi. Clearly, more material is needed before isolated M1s and M2s can be differentiated. In order to provide a full description of the dentition of Eoconodon nidhoggi, the following descriptions, illustrations (Figure 2.4-2.9), and dimensions (Table 1) of its lower molars are reproduced from Clemens and Williamson (2005, p. 210) with emendations to account for subsequently discovered material and differences in journal format. Three fragmentary m1s are referable to Eoconodon nidhoggi. Their protoconids and slightly lower metaconids are separate cusps only in the upper one-third of the trigonid. The paraconid, distinctly lower and projecting anteriorly, is situated in front of the base of the notch in the protocristid. Differing from m1s of E. gaudrianus and the new species described below, the labial basal cingulid is continuous along the entire length of the tooth. The hypoconid is the largest and highest talonid cusp. Hypoconulid and entoconid are subequal in height and closely approximated to form a transverse ridge. The entocristid extends to the back of the trigonid but decreases in height anteriorly so the trough-like talonid basin opens lingually behind the metaconid. The two roots are of approximately equal size. Morphologically m1s and m2s of Eoconodon nidhoggi only differ slightly in that the paraconid of m2 is less salient anteriorly and situated slightly labial to the metaconid, not in front of the notch in the protocristid (Figure 2.4-2.6). The labial basal cingulid is not as strongly developed as it is on m1. The last molar is known from the type specimen of Eoconodon nidhoggi (UMVP 1471) and two other isolated teeth (Figure 2.7-2.9). Comparisons of isolated m2s and m3s suggest that in any individual m3 might have been slightly longer (Table 1). In comparison to m2s the trigonid of m3 is slightly lower relative to the height of the talonid. The protoconid is more robust and slightly higher than the metaconid. On m3s the paralophid extends anterolingually down the protoconid and then turns to reach the small paraconid. On the type the labial basal cingulid is interrupted on the posterolabial side of the trigonid. On the referred molars this cingulid begins below the paraconid and continues as a narrow shelf across the trigonid and talonid before rising to end at the base of the hypoconulid. As on other molars, the hypoconid of m3 is the largest talonid cusp and linked to the slightly lower hypoconulid by a strong ridge. The entoconid is distinctly smaller and separated by a groove from the hypoconulid. Extending anteriorly from the entoconid, the entocristid decreases in height and ends at the lingual side of the base of the metaconid. Just posterior to the metaconid the talonid basin is broadly open lingually across the low entocristid. Of the two roots, the posterior is significantly larger and directed posteroventrally. Unlike m3s of E. gaudrianus from the San Juan Basin, the back of the hypoconulid is more vertically oriented and separated from the root by a distinct notch. Eoconodon hutchisoni new species Holotype. UCMP 156108, a left M2 preserved in a fragment of maxillary. Type Locality. UCMP V81028, Tullock Formation, Garfield County, northeastern Montana, Pu2/Pu3. Hypodigm and localities. ?DP3 or DP4, UCMP 145677 (V-73080); P4, LACM 28140 (V-72125), UCMP 170844 (V-73080), UCMP 156102, 156118 (V-73082); M1, UCMP 156100, 156104 (V-73080), UCMP 156120 (V-73096); M2, UMVP 1450 (V-71202), UCMP 156103 (V-72130), UCMP 156105, 156106, 170845, 186684 (V-73080), UCMP 156108 (V-81028); M3, UCMP 156109 (V-72130), UCMP 170846 (V-73080), UCMP 156119 (V-74123); p4, UCMP 145676, 156112, 156113, 156114, 156115, 170869 (V-73080); m1, UCMP 145324 (V-72130), UCMP 110515, 192011, 192012 (V-73080), UCMP 110590, 192016 (V-73096); m2, LACM 28138 (V-72125), UCMP 110516, 145326, 145327, 145333, 192013 (V-73080), UCMP 156117 (V-75196); m3, LACM 28139 (V-72125), UCMP 156116 (V-72131), UCMP 145331 (V-73080), UCMP 192017 (V99438); casts of two fragments of a dentary, one containing m2-3, the other preserving part of the symphyseal region, UCMP 124393 (V-80002). All of the following UCMP localities are in Garbani Channel deposits either at the main quarry locality or elsewhere in the western headwaters of Cottonwood Creek (see Archibald 1982): V-72125, V-72130, V-72131, V-73080, V-73082, V-73096, V-74123, V-81028, and V-99438. Additional specimens of E. hutchisoni have been found at Purgatory Hill (UCMP loc. V-71202), Kerr Butte (UCMP loc. V-75196, see Archibald 1982, p. 33-34), and a locality somewhere in the valley of Hell Creek (V-80002) discovered by Reverend Wayne Moore. Etymology. Named to recognize the many contributions to vertebrate paleontology made by Dr. J. Howard Hutchison. The type specimen is just one of a multitude of fossils that he discovered and that significantly add to the basis of research on the faunas of the Hell Creek and Tullock Formations of northeastern Montana. Diagnosis. Postcanine dentition of Eoconodon hutchisoni, where known, is smaller than that of E. coryphaeus or the observed ranges of variation slightly overlap (Table 1). Its postcanine dentition is larger than those of E. nidhoggi and E. ginibitohia but again a few observed ranges of variation slightly overlap. With the exception of the distinctly smaller p4s and m3s of E. hutchisoni, the other lower postcanines in the small sample of E. gaudrianus are either slightly smaller than or within the lower end of the observed range of variation of those of E. hutchisoni. The P4 of Eoconodon hutchisoni differs from that of E. gaudrianus and the P4 tentatively referred to E. nidhoggi (UMVP 1475) in its significantly larger protocone that, particularly in comparison to E. gaudrianus, is more distinctly separated from the base of the paracone. P4s of E. hutchisoni have a small, metacone-like expansion on the posterior slope of the paracone, which is lacking in E. gaudrianus, and possibly E. nidhoggi, and the posterior margin of the ectocingulum is more rugose. Differing from M1s or M2s of E. nidhoggi in which the metacone is slightly smaller than the paracone, on M1s of E. hutchisoni these cusps are subequal while on M2s the metacone is the smaller cusp. On M1s of E. hutchisoni the postcingulum is broader than that of the M1s or M2s of E. nidhoggi and has a distinct hypocone. The metacone of M3 is much smaller than the paracone. A metaconule is present but the paraconule is either small or missing. Its trigon basin is divided by a low transverse crest. Precingulum and postcingulum are narrow and do not meet on the lingual face of the protocone. The protoconids of p4s of Eoconodon hutchisoni are not as distinctly canted posteriorly as those of E. gaudrianus, and, particularly, E. coryphaeus. The lengths of the m1s and m2s of E. hutchisoni are smaller than those of E. coryphaeus, greater than those of E. nidhoggi and E. ginibitohia, and overlap with those of E. gaudrianus (Table 1). In contrast to the other currently recognized species, the m3 of E. hutchisoni is distinctly shorter and narrower than m2. (Based on means, see Table 1, the crown length of m3 is 86% of that of m2 and the trigonid width of m3 is 84% of that of m2.) The m3 talonid is reduced in size relative to the trigonid, and on one specimen the talonid cusps are reduced to crenulations around the talonid basin. On m3s of E. hutchisoni the posterior slope of the hypoconulid is separated from the posterior root by a distinct notch, while in E. gaudrianus (e.g., AMNH 3400 and the fragmentary m3 of AMNH 58116) the posterior slope of the hypoconulid merges smoothly with the surface of the enlarged posterior root. Description and comparisons. The majority of the specimens forming the available sample of Eoconodon hutchisoni are isolated teeth. One of two exceptions is UCMP 156108, the type specimen, which consists of an M2 preserved in a fragment of maxillary including the posterior walls of the alveoli of M1, the anterior walls of alveoli of M3, and the anterior base of the zygomatic arch. The other is UCMP 124393, which consists of two, very probably associated, fragments of a dentary. One contains both m2 and m3. The other preserves the symphyseal region. Identifications of the positions of the postcanines of E. hutchisoni not represented in these specimens are based on comparisons with the dentitions of other species of Eoconodon, primarily E. coryphaeus.
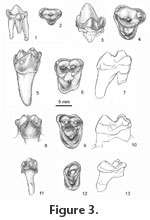
In occlusal view, the parastyle of the P4 is large, rounded, and anteriorly projecting (Figure 3.3-3.4). Viewed laterally, the anterior crest of the high paracone is longer than the posterior, which carries a small metacone-like expansion and terminates in a small posterior accessory cusp. Above the apex of the paracone, the ectocingulum narrows to a faint ridge. Posteriorly it expands onto the anteroposteriorly short but broad metastylar region. The prominent protocone is linked to the paracone by a low, rounded surface delimited anteriorly and posteriorly by traces of cingula extending from the protocone to the parastylar and metastylar regions. Conules are lacking. A small postcingulum is present on one (LACM 28140) of the three specimens preserving this region. The three roots are of approximately equal size and not divergent. The salient, rounded parastylar region of M1 carries a small, distinct cusp (Figure 3.5-3.7). A very shallow ectoflexus divides the ectocingulum into symmetrical lobes. Usually a small mesostylar cusp is present. Paracone and metacone are low, conical cusps of essentially equal size. Low ridges extend from the paraconule and metaconule toward the bases of the paracone and metacone. A preparaconule crista extends to the parastyle; the postmetaconule crista extends to the posterior labial corner of the crown. The areas between the conules, paracone, and metacone delimited by these crests are essentially flat surfaces. The labial side of the high protocone is also relatively flat. The trigon is essentially an antero-posteriorly oriented trough, not a basin. It consists of the two flat areas labial to the conules and the flat but more dorsally placed labial surface of the protocone. A narrow precingulum is present. The posterior slope of the protocone is inflated, and bears a postcingulum that terminates lingually in a small hypocone. On M1s and other upper molars irregular bumps and nearly vertical striae can be present particularly on the near vertical, lingual surfaces of the cusps. Of the three roots, the lingual is much greater in length and diameter at base of crown than the other two. The M2s are more transversely broadened than M1s, which is reflected in their length/width ratios (M2, 0.72-0.76 vs. M1, 0.83). In occlusal view (Figure 3.8-3.10), the labial margin of the ectocingulum is asymmetrical with the posterior lobe slightly deflected anteriorly. Unlike M1s the paracone of M2s is somewhat larger than the metacone. Also the trigon is shallowly but distinctly basined. Differing from M1s, the precingulum and postcingulum are of approximately equal width and lack a hypocone. These cingula usually do not meet across the labial face of the protocone, but on two of the six teeth there is a narrow connection. Of the three roots, the lingual is largest and longest. On the basis of their large size and bluntly conical cusps three isolated M3s are referred to Eoconodon hutchisoni. On these teeth the ectocingulum extends around the not greatly expanded parastylar region (Figure 3.11- 3.13). A very small parastylar cusp is situated lingually and connected to the paracone by a low crest. Posterolabially, the ectocingulum is continuous; its crest is crenulated but distinct stylar cusps are not developed. The paracone is much larger than the metacone. A small metaconule is present. On one tooth a smaller paraconule is present; the others lack this cusp. The morphology of the trigon varies. On one tooth (UCMP 156109), a distinct transverse ridge divides the trigon into anterior and posterior basins. Its precingulum and postcingulum are well developed but do not meet lingually. On the other referred teeth the trigon basins are smaller. Their occlusal surfaces are slightly convex but lack a pronounced transverse crest. The precingulum and postcingulum are short and poorly defined. The crown was supported by two roots; the lingual is slightly larger.
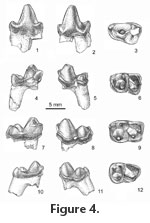
The protoconid and slightly lower metaconid of m1 are separate cusps only in approximately the upper one-third of the trigonid (Figure 4.4-4.6). The paraconid is distinctly lower, is situated in front of the notch separating the protoconid and metaconid, and projects anteriorly. A broad anterior basal cingulid extends ventrolabially from the base of the paraconid to the base of the trigonid, narrows or is interrupted across the labial face of the trigonid, continues posteriorly across the labial face of the talonid, and then turns dorsally to end at the base of the hypoconulid. The hypoconid is the largest and highest talonid cusp. Hypoconulid and entoconid are subequal in height and closely approximated to form a transverse ridge (postcristid). From the entoconid, the entocristid decreases in height toward the trigonid allowing the trough-like talonid basin to open lingually behind the metaconid. The two roots are of approximately equal size. The m2s differ slightly from m1s in that the paraconid is not as salient anteriorly, usually is more lingually situated, and the apices of paraconid and metaconid are more closely approximated (Figure 4.7-4.9). In Eoconodon coryphaeus, whose lower dentition is known from a large number of specimens (Table 1), the length of m1 tends to be slightly shorter than that of m2. Also, the width of the m1 trigonid is less than that of the talonid, while on m2 the trigonid is slightly wider than the talonid. These proportions, which describe the m2 preserved in a fragment of dentary (UCMP 124343), have been used to distinguish isolated m1s and m2s of E. hutchisoni. The protoconid of m3 is more robust and slightly higher than the metaconid; the paraconid is distinctly smaller and, unlike m1 and m2, situated directly in front of the metaconid (Figure 4.10-4.13). The talonids of most m3s resemble those of the other molars in that the hypoconid is the largest talonid cusp, and the slightly smaller hypoconulid is linked to the entoconid by a strong ridge (postcristid). On one tooth (UCMP 192017) the talonid basin is rimmed by a series of small cusps whose homologies are not apparent. The talonid is much narrower than the trigonid, although the degree of its diminution relative to the trigonid is variable. The labial base of the crown is expanded, and on some teeth a cingulid is present on the anterior face of the trigonid and labial side of the talonid. Of the two roots the posterior is separated from the crown by a shallow notch around the top of the root, significantly larger, and directed posteroventrally,
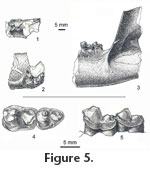
One fragment of UCMP 124393 includes parts of the symphyseal region, the alveolus for the canine, and alveoli for two, two-rooted premolars (Figure 5.1-5.2). Depth of the dentary below the premolars is ca. 2.8 cm, which is comparable to that of the posterior fragment. The positions of the premolar alveoli suggest the anterior two premolars were crowded and set en echelon with their posterior roots lingual to the anterior. There is no evidence of mental foramina on what remains of the labial side of the dentary. Medially, what remains of its rugose surface indicates the symphysis was massive. The posterior edge of the symphysis is rounded and, ventrally, projects posteriorly. From this projection a low but distinct ridge extends posteriorly, parallel to the bottom of the dentary. The second fragment consists of part of the posterior end of the dentary including the nearly vertical anterior edge of the coronoid process as well as m2 and m3 (Figure 5.3). The masseteric fossa is sharply delimited anteriorly. On the lingual side of the dentary a horizontal ridge is present ventral to the coronoid process and ca. 8 mm below the alveolar level of the ramus. Two fragmentary dentaries of Eoconodon gaudrianus (AMNH 3400 and 58116) are smaller than but resemble UCMP 124393 in their similar depth throughout their length and heavily built symphyses. Dentaries of E. coryphaeus exhibit a similar massive structure. On most specimens of E. coryphaeus the anterior part of the dentary is absent or poorly preserved. Where it is present usually there is no evidence of crowding of the premolars. In contrast, the anterior part of the dentary of AMNH 16333 preserves the root of the canine and parts of the crowns of p1-2 and the complete p4 as well as m1-2. The large canine is separated from p1 by a short (ca. 4.0 mm) diastema, which is the only significant gap in the postcanine dentition. The damaged crown of p1 had a smaller diameter than its single root. A thin interalveolar wall, ca. 0.5 mm, separates the roots of p1 and p2. Three roots support p2, the larger anterior and posterior roots and a smaller, labial root below the apex of the main cusp. The p2 and p3 are set en echelon with p3 overlapping the labial side of p2 by ca. 2 mm, which is the only observed occurrence of crowding in the premolar dentition of E. coryphaeus. In an early study of the enamel microstructure of Paleocene mammals, Koenigswald et al. (1987) described the structural complexity of the enamel of Eoconodon coryphaeus (identified as E. heilprinianus) and E. gaudrianus. They reported organization of the enamel prisms into groups following divergent paths outward from the enamel-dentine junction creating Hunter-Schreger bands. When a tooth is viewed laterally or in tangential section primitively Hunter-Schreger bands are approximately parallel and relatively straight. In more derived structural patterns the bands are undulating culminating in a zig-zag pattern, which increases the resistance of the enamel to splitting (see Stefen 1997a, 1997b). Stefen (1999) examined sections through the trigon of UC 156100, now identified as an M1 of E. hutchisoni, and a fragment of another upper molar, UCMP 171513, of this species. In these specimens the Hunter-Schreger bands exhibit an increasing degree of waviness from the base toward the apex of the crown. Currently species of Eoconodon provide the earliest record of this derived microstructural pattern in eutherian mammals (Stefen 1999). Comments. Determination of the generic reference of E. hutchisoni highlights the current limitations of the definitions and diagnoses of Eoconodon and Goniacodon, which is known from Pu3 and Torrejonian faunas in the Western Interior, primarily the San Juan Basin, New Mexico, and Dragon Canyon, Utah (Lofgren et al. 2004). Matthew (1937) reviewed Goniacodon and its type species, G. levisanus, and cited several dental and mandibular characters distinguishing it from Eoconodon. Upper molars are less quadrate – considerably wider than long – than those of Eoconodon. M1 is as large as or larger than M2; the proportions of these molars in Eoconodon are reversed. Protoconules are absent but metaconules are distinct. In Goniacodon, hypocones are present on M1-2. Paraconids of the lower molars are relatively smaller than those of the molars of Eoconodon. Protoconid of p4 of Goniacodon is "...rather recurved towards the tip than backwardly pitched from the base (Matthew 1937, p. 88)" as is the case in Eoconodon. The mandibular symphysis of Goniacodon was described as "...ovate and widest behind extending back to beneath the third premolar while in Eoconodon it is widest anteriorly and pointed behind" (Matthew 1937 p. 86). In Goniacodon, the mental foramina are positioned far forward below p1 and p2. The M3 and m3 were described as "reduced" (Matthew 1937, p. 86) or "much reduced" (Matthew 1937, p. 87) in comparison to the second molars. As is the case with molar proportions, quantitative data were not given. In almost all of the characters cited by Matthew (1937) E. hutchisoni resembles other species of Eoconodon and not Goniacodon, which appears to have more derived character states. An exception is the reduction in relative size of m3 of E. hutchisoni (M3 of this species has yet to be identified). In the fragmentary mandible of E. hutchisoni (UCMP 124393) the length of m3 is 83% of that of m2; trigonid width of m3 is 82% of that of m2. Adding isolated teeth, the mean values of length and trigonid widths of m3s are, respectively, 86% and 84% of those of m2s. Where known, the m3s of other species of Eoconodon are approximately the same size as their m2s. Dimensions of m2 and m3 of G. levisanus, supplied by Thomas Williamson (personal commun., 2007), indicate that the mean length of m3s in this species is approximately 70% of the mean length of m2s, and mean trigonid width of m3s is 68% of that of m2s. Although the length and trigonid width of m3 relative to those of m2 of E. hutchisoni are smaller than in other species of Eoconodon, they are not as greatly reduced as in Goniacodon levisanus. Eoconodon copanus Van Valen, 1978 Type. PU 13290, left m1? (Jepsen 1930, plate IV, fig. 5) Type locality. Mantua Lentil, Polecat Bench Formation, Wyoming (Jepsen 1930; Van Valen 1978) Diagnosis. "Lower molars elongate, entoconid small and distal. Same as Eoconodon sp. of Jepsen (1930, Plate IV, fig. 5) M?1 l, 10.4, trd w, 5.6, tald w, 5.4 (Van Valen 1978, p. 57)" . Comments. The type and only specimen of Eoconodon copanus (PU 13290) is from the sample of the early Puercan (Pu1) Mantua local fauna. Its salient paraconid suggests that the tooth is an m1. A fracture extends diagonally along the floor of the trough-like, lingually-sloping talonid basin. The part of the talonid containing both the entoconid and hypoconulid is shifted slightly distally. Some separation of the fragments has occurred and could influence the measurements of length of the crown and width of the talonid. Dimensions measured on a cast of PU 13290 (Table 1) differ somewhat from those reported by Van Valen (1978). Depending upon which set of measurements of PU 13290 is used, its crown length falls just within the lower limit of the observed range of variation of Eoconodon coryphaeus or between that and the observed range of variation in the smaller E. hutchisoni. Both of the measurements of its trigonid width are distinctly smaller than those of E. coryphaeus. The measurement reported by Van Valen (1978) falls just within the lower bound of the observed range of variation of E. hutchisoni; the measurement based on the cast is smaller. Both sets of measurements indicate that the talonid of PU 13290 is narrower than the talonids of m1s of E. coryphaeus and E. hutchisoni. This relative narrowness of the crown of PU 13290 reflects the lack of expansion of the base of the crown and development of distinct basal cingulids that characterize the m1s and m2s of E. hutchisoni and E. coryphaeus. Van Valen (1978) cited the small size and distal position of the entoconid in his diagnosis of Eoconodon copanus. The entoconid is small but, in comparison to the talonid cusps of other species of Eoconodon, it is not particularly reduced, and its position is not greatly different from that in E. nidhoggi and E. hutchisoni. Until a larger sample is discovered, the relative narrowness of the crown and its length – approximately intermediate between those of m1s of E. nidhoggi and E. hutchisoni – warrant continued recognition of E. copanus as a distinct species. Eoconodon sp. Numerous fossil localities in a remarkably thick (over 530 m, Eberle and Lillegraven 1998a) and structurally uncomplicated section of the Ferris Formation in the Hanna Basin, south central Wyoming, have yielded remains of Lancian and Puercan (Pu1 through Pu3) mammals. Eberle and Lillegraven (1998b) documented the presence of Eoconodon in the Pu1 and Pu2 interval zones. The Pu1 record is based on an isolated right upper molar (UW 26187), probably an M2. Eberle and Lillegraven (1998b, p. 102) noted that this molar is about half the size of homologous molars of E. coryphaeus and somewhat smaller than those of E. gaudrianus and "...the unnamed larger species from Purgatory Hill..." (=E. hutchisoni). Basing an estimate of the sizes of its upper molars on the dimensions of the type of E. nidhoggi, an m3, they suggested that UW 26187 probably was smaller than the upper molars of E. nidhoggi. Although few in number, the sample of M1s or M2s of E. nidhoggi described above fully supports their estimate. Also, the molars of E. nidhoggi differ in the similar length and width of their precingulum and postcingulum and the absence of a distinct hypocone. Again based on an estimate of size of the upper molars of E. copanus, Eberle and Lillegraven (1998b) argued that UW 26187 probably is too small to be referable to this species, the only Pu1 species of Eoconodon currently recognized and named. The second specimen of Eoconodon from the Hanna Basin, found at a Pu2 locality, is a fragment of the labial part of an upper molar (UW 26221), which, as far as can be determined, resembles UW 26187. Eberle and Lillegraven (1998b) suggested that the two specimens from the Ferris Formation represent a new species of Eoconodon. With discovery of larger samples, it would be appropriate to compare Eoconodon sp. from the Ferris Basin with E. ginibitohia (Clemens and Williamson 2005) from the San Juan Basin, which is the smallest, currently named species of the genus. Although small, the records of Eoconodon from the Ferris Formation are significant in documenting the presence of a second species of the genus in Pu1 faunas of the Western Interior. Although the collections from Pu3 localities are described as quite large consisting of "hundreds" of mammalian specimens (Eberle and Lillegraven 1998b, p. 110), a record of Eoconodon or another "triisodontid" is lacking. |
|