|
|
|
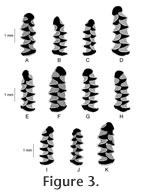
RESULTSThe four arvicoline rodent taxa identified from SCC include extinct (Microtus meadensis, Microtus paroperarius) and extant voles [Microtus sp. (not Microtus meadensis or Microtus paroperarius), Lemmiscus curtatus]. Both Microtus sp. and Lemmiscus curtatus are characterized in the fauna by multiple morphotypes. These include the four-triangle form of Lemmiscus curtatus, a morphotype that persisted only into the early Holocene (Barnosky and Bell 2003; Bell and Jass 2004). Identifying characteristics and abundance of each identified taxon are discussed below. Two hundred thirty-nine specimens (80.7%) were identified as Microtus sp., making it the dominant taxon in the SCC deposit. These specimens exhibit greater labial-lingual length in T1 than T2, a character that can be used to distinguish Microtus from Lemmiscus (Barnosky and Rasmussen 1998). Specimens identified as Microtus sp. in this paper are characterized by the presence of at least five closed, alternating triangles, distinguishing them from both Microtus paroperarius and Microtus meadensis. Previous reports on the SCC fauna separated Microtus cf. M. montanus (Goodrich 1965; Miller 1979) and Microtus cf. M. longicaudus (Mead et al. 1982) as distinct from other specimens referred to Microtus sp. Given the current state of knowledge regarding identification of m1s of Microtus at the species level (see Bell et al. 2004a for discussion), there is no morphological basis for the identification of Microtus cf. M. montanus and Microtus cf. M. longicaudus from SCC. I consider previous reports of these taxa to be synonymous with Microtus sp. as reported here.
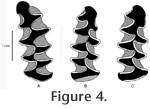
Three of the 296 specimens (1.0%) were identified as Microtus paroperarius. These m1s have four closed, alternating triangles, followed by a well-developed fifth triangle that is variably confluent with the secondary wings and anterior cap (Figure 4B, C). Two of these specimens exhibit a state where the fifth triangle is pinched from the secondary wings (e.g., Figure 4B). In one specimen (SCCAR-212), the fifth triangle is broadly confluent with the secondary wings (Figure 4C). This variation is consistent with that seen in other samples of Microtus paroperarius, but is also known to be similar or identical to the living Microtus oeconomus (van der Meulen 1978). Additionally, this morphology is known to occur in very low percentages in some other extant species of Microtus (Bell and Repenning 1999).
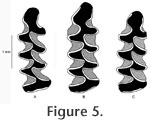
Four-triangle forms of Lemmiscus curtatus are noteworthy because that morphology is not known to occur in extant populations of sagebrush voles (Bell and Barnosky 2000; Barnosky and Bell 2003), and the loss of this morphotype may represent one of the few small mammal 'extinction' events near the early Holocene (Bell and Jass 2004). Smith Creek Cave represents one of the few localities known to preserve specimens of Lemmiscus curtatus that have a four-triangle morphology (other localities were reviewed by Bell and Jass 2004). Given the fact that three of the localities containing a four-triangle form of Lemmiscus curtatus occur in, or near Smith Creek Canyon, it seems a strong possibility that additional Pleistocene localities in the vicinity (e.g., Crystal Ball Cave) may also preserve this morphotype. In addition to the specimens reported here, two other arvicoline rodent taxa were reported from SCC. A single specimen (NAUQSP 17910) of Phenacomys that was collected from a back-dirt pile in SCC is housed at the Laboratory of Quaternary Paleontology, Northern Arizona University. The specimen was not examined for this study but was described and illustrated by Mead et al. (1982) as Phenacomys cf. P. intermedius and by Repenning and Grady (1988) as Phenacomys albipes. Specimens of Mictomys borealis were reported from both Cathedral Cave and Smith Creek Cave (Mead et al. 1992). The record from Cathedral Cave consists of multiple specimens, but only a single M2 was reported from SCC. A subsequent re-examination of arvicoline rodents from Cathedral Cave (Bell 1995) revised the identification of Mictomys borealis but did not address the specimen from SCC. During the course of this study, no additional specimens of Mictomys were found in the SCC material housed at the Nevada State Museum. In the absence of additional specimens of Mictomys from SCC (e.g., m1s) that would allow for a more confident taxonomic placement, I consider the single record reported by Mead et al. (1992) to be Mictomys sp. |
|